Download presentation
Presentation is loading. Please wait.

2
Dynamics of Networks 2 Synchrony & Balanced Colourings Ian Stewart Mathematics Institute University of Warwick Ian Stewart Mathematics Institute University of Warwick UK-Japan Winter School Dynamics and Complexity

3
Network A network or directed graph consists of a set of: consists of a set of: nodes or vertices or cellsnodes or vertices or cells connected by directed edges or arrowsdirected edges or arrows

4
dx 1 /dt = f(x 1,x 2,x 4,x 5 ) dx 2 /dt = f(x 2,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 4 ) dx 4 /dt = g(x 4,x 2,x 4 ) dx 5 /dt = h(x 5,x 4 ) Admissible ODEs
 dx 2 /dt = f(x 2,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 4 ) dx 4 /dt = g(x 4,x 2,x 4 ) dx 5 /dt = h(x 5,x 4 ) Admissible ODEs")
5
Pattern of Synchrony Cells c, d are synchronous on some trajectory x(t) if x c (t) = x d (t) for all t Defines an equivalence relation ~ for which c ~ d if and only if c, d are synchronous Call ~ a pattern of synchrony
 if x c (t) = x d (t) for all t Defines an equivalence relation ~ for which c ~ d if and only if c, d are synchronous Call ~ a pattern of synchrony")
6
Colouring Interpretation More intuitively, colour cells with different colours, so that cells have the same colour if and only if they are synchronous Synchrony Space or Polydiagonal ~ = {x : x c = x d whenever c ~ d} ~ = {x : x c = x d whenever c ~ d} This is the set of all cell states with the pattern of synchrony ~ It forms a vector subspace of phase space P

7
dx 1 /dt = f(x 1,x 2,x 4,x 5 ) dx 2 /dt = f(x 2,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 4 ) dx 4 /dt = g(x 4,x 2,x 4 ) dx 5 /dt = h(x 5,x 4 )
 dx 2 /dt = f(x 2,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 4 ) dx 4 /dt = g(x 4,x 2,x 4 ) dx 5 /dt = h(x 5,x 4 )")
8
dx 1 /dt = f(x 1,x 1,x 3,x 5 ) dx 1 /dt = f(x 2,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 3 ) dx 5 /dt = h(x 5,x 3 ) x 1 =x 2 x 3 =x 4
 dx 1 /dt = f(x 2,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 3 ) dx 5 /dt = h(x 5,x 3 ) x 1 =x 2 x 3 =x 4")
9
dx 1 /dt = f(x 1,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 3 ) dx 5 /dt = h(x 5,x 3 ) restricted equations
 dx 3 /dt = g(x 3,x 1,x 3 ) dx 5 /dt = h(x 5,x 3 ) restricted equations")
10
dx 1 /dt = f(x 1,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 3 ) dx 5 /dt = h(x 5,x 3 ) restricted to synchrony space {(x 1,x 1,x 3,x 3,x 5 )}
 dx 3 /dt = g(x 3,x 1,x 3 ) dx 5 /dt = h(x 5,x 3 ) restricted to synchrony space {(x 1,x 1,x 3,x 3,x 5 )}")
11
dx 1 /dt = f(x 1,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 3 ) dx 5 /dt = h(x 5,x 3 ) restricted to synchrony space {(x 1,x 1,x 3,x 3,x 5 )} quotient network
 dx 3 /dt = g(x 3,x 1,x 3 ) dx 5 /dt = h(x 5,x 3 ) restricted to synchrony space {(x 1,x 1,x 3,x 3,x 5 )} quotient network")
12
States on the synchrony space satisfy the restricted equations and have the pattern of synchrony determined by the colouring The dynamics of these states is determined by the quotient network

13
A pattern of synchrony ~ is robust if ~ is invariant under all admissible vector fields That is: any initial condition with that pattern continues to have that pattern for all time Robust Synchrony

14
A pattern of synchrony ~ (or the associated colouring) is balanced if cells with the same colour have input sets with the same colours (of tail cells) up to some type-preserving permutation (input isomorphism) Theorem A pattern of synchrony ~ is robust if and only if it is balanced Balanced Patterns and Colourings
 is balanced if cells with the same colour have input sets with the same colours (of tail cells) up to some type-preserving permutation (input isomorphism) Theorem A pattern of synchrony ~ is robust if and only if it is balanced Balanced Patterns and Colourings")
16
Balanced colouring: cells with same colour have input sets with same colours

17
This colouring is not balanced

18
Nearest-neighbour coupling Balanced 2-colouring Examplethe Square Lattice

19
For this pattern, interchanging black and white along any upward sloping diagonal leads to another balanced 2-colouring Examplethe Square Lattice

20
The Origin of Species

21
Allopatric speciation Geographical or other barrier prevents gene-flow EasilyunderstoodandWYSIWYG The Origin of Species

22
Sympatric speciation No such barrier; species split in the same location even with panmixis Sympatric speciation appears paradoxical, but can be viewed as a form of symmetry-breaking The Origin of Species

23
Darwins Finches Galápagos Islands

24
Darwins Finches

25
1. Large cactus finch (Geospiza conirostris) 2. Large ground finch (Geospiza magnirostris) 3. Medium ground finch (Geospiza fortis) 4. Cactus finch (Geospiza scandens) 5. Sharp-beaked ground finch (Geospiza difficilis) 6. Small ground finch (Geospiza fuliginosa) 7. Woodpecker finch (Cactospiza pallida) 8. Vegetarian tree finch (Platyspiza crassirostris) 9. Medium tree finch (Camarhynchus pauper) 10. Large tree finch (Camarhynchus psittacula) 11. Small tree finch (Camarhynchus parvulus) 12. Warbler finch (Certhidia olivacea) 13. Mangrove finch (Cactospiza heliobates) Darwins Finches
 4. Cactus finch (Geospiza scandens) 5. Sharp-beaked ground finch (Geospiza difficilis) 6. Small ground finch (Geospiza fuliginosa) 7. Woodpecker finch (Cactospiza pallida) 8. Vegetarian tree finch (Platyspiza crassirostris) 9. Medium tree finch (Camarhynchus pauper) 10. Large tree finch (Camarhynchus psittacula) 11. Small tree finch (Camarhynchus parvulus) 12. Warbler finch (Certhidia olivacea) 13. Mangrove finch (Cactospiza heliobates) Darwins Finches.")
26
1. Large cactus finch (Geospiza conirostris) 2. Large ground finch (Geospiza magnirostris) 3. Medium ground finch (Geospiza fortis) 4. Cactus finch (Geospiza scandens) 5. Sharp-beaked ground finch (Geospiza difficilis) 6. Small ground finch (Geospiza fuliginosa) 7. Woodpecker finch (Cactospiza pallida) 8. Vegetarian tree finch (Platyspiza crassirostris) 9. Medium tree finch (Camarhynchus pauper) 10. Large tree finch (Camarhynchus psittacula) 11. Small tree finch (Camarhynchus parvulus) 12. Warbler finch (Certhidia olivacea) 13. Mangrove finch (Cactospiza heliobates) Darwins Finches
 4. Cactus finch (Geospiza scandens) 5. Sharp-beaked ground finch (Geospiza difficilis) 6. Small ground finch (Geospiza fuliginosa) 7. Woodpecker finch (Cactospiza pallida) 8. Vegetarian tree finch (Platyspiza crassirostris) 9. Medium tree finch (Camarhynchus pauper) 10. Large tree finch (Camarhynchus psittacula) 11. Small tree finch (Camarhynchus parvulus) 12. Warbler finch (Certhidia olivacea) 13. Mangrove finch (Cactospiza heliobates) Darwins Finches.")
28
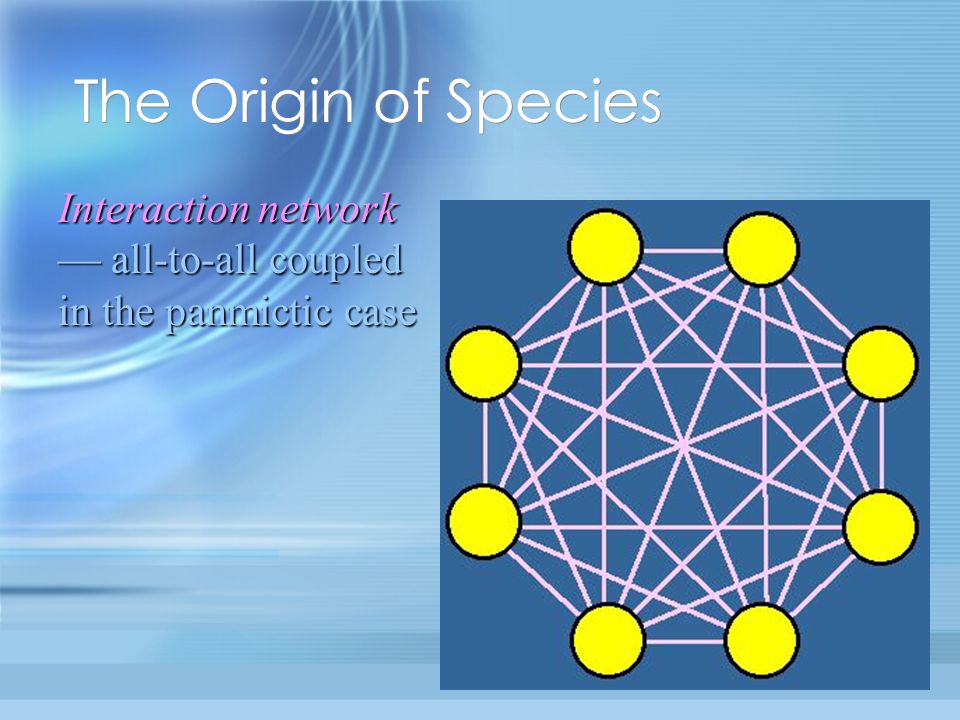
Interaction network all-to-all coupled in the panmictic case The Origin of Species

29
With generic nonlinear dynamics, instability of the single-species state leads to symmetry-breaking bifurcation. Universal features are: Split is a jump bifurcation Mean phenotypes stay constant Usual split is to two species The Origin of Species

30
Simulation with 50 nodes The Origin of Species

31
Geospiza fuliginosa Geospiza fortis Albemarle/Isabela Daphne Crossman/ Los Hermanos Beak size in millimetres Character displacement Not speciation, but might be OK as a surrogate Darwins Finches

32
The Origin of Species

33
J.Cohen and I.Stewart. Polymorphism viewed as phenotypic symmetry-breaking, in Nonlinear Phenomena in Biological and Physical Sciences (eds. S.K.Malik, M.K.Chandra-sekharan, and N.Pradhan), Indian National Science Academy, New Delhi 2000, 1-63 I.Stewart. Self-organization in evolution: a mathematical perspective, Nobel Symposium Proceedings, Phil. Trans. Roy. Soc. Lond. A 361 (2003) 1101-1123. I.Stewart., T.Elmhirst and J.Cohen. Symmetry-breaking as an origin of species, in Bifurcations, Symmetry, and Patterns (eds. J. Buescu, S. Castro, A.P.S. Dias, and I. Labouriau), Birkhäuser, Basel 2003, 3-54. I.Stewart. Speciation: a case study in symmetric bifurcation theory, Univ. Iagellonicae Acta Math. 41 (2003) 67-88. J.Cohen and I.Stewart. Polymorphism viewed as phenotypic symmetry-breaking, in Nonlinear Phenomena in Biological and Physical Sciences (eds. S.K.Malik, M.K.Chandra-sekharan, and N.Pradhan), Indian National Science Academy, New Delhi 2000, 1-63 I.Stewart. Self-organization in evolution: a mathematical perspective, Nobel Symposium Proceedings, Phil. Trans. Roy. Soc. Lond. A 361 (2003) 1101-1123. I.Stewart., T.Elmhirst and J.Cohen. Symmetry-breaking as an origin of species, in Bifurcations, Symmetry, and Patterns (eds. J. Buescu, S. Castro, A.P.S. Dias, and I. Labouriau), Birkhäuser, Basel 2003, 3-54. I.Stewart. Speciation: a case study in symmetric bifurcation theory, Univ. Iagellonicae Acta Math. 41 (2003) 67-88.
, Indian National Science Academy, New Delhi 2000, 1-63 I.Stewart. Self-organization in evolution: a mathematical perspective, Nobel Symposium Proceedings, Phil. Trans. Roy. Soc. Lond. A 361 (2003) I.Stewart., T.Elmhirst and J.Cohen. Symmetry-breaking as an origin of species, in Bifurcations, Symmetry, and Patterns (eds. J. Buescu, S. Castro, A.P.S. Dias, and I. Labouriau), Birkhäuser, Basel 2003, I.Stewart. Speciation: a case study in symmetric bifurcation theory, Univ. Iagellonicae Acta Math. 41 (2003) J.Cohen and I.Stewart. Polymorphism viewed as phenotypic symmetry-breaking, in Nonlinear Phenomena in Biological and Physical Sciences (eds. S.K.Malik, M.K.Chandra-sekharan, and N.Pradhan), Indian National Science Academy, New Delhi 2000, 1-63 I.Stewart. Self-organization in evolution: a mathematical perspective, Nobel Symposium Proceedings, Phil. Trans. Roy. Soc. Lond. A 361 (2003) I.Stewart., T.Elmhirst and J.Cohen. Symmetry-breaking as an origin of species, in Bifurcations, Symmetry, and Patterns (eds. J. Buescu, S. Castro, A.P.S. Dias, and I. Labouriau), Birkhäuser, Basel 2003, I.Stewart. Speciation: a case study in symmetric bifurcation theory, Univ. Iagellonicae Acta Math. 41 (2003)")
34
Equilibrium States x*is an equilibrium state for an admissible f if and only if f(x*) = 0 Associated Pattern of Synchrony Use the cell coordinates of x* to define a colouring: then c and d have the same colour if and only if x* c = x* d Determines an equivalence relation ~ x* Rigid Equilibria
 = 0 Associated Pattern of Synchrony Use the cell coordinates of x* to define a colouring: then c and d have the same colour if and only if x* c = x* d Determines an equivalence relation ~ x* Rigid Equilibria")
35
An equilibrium x* is hyperbolic if the Jacobian Df| x* has no eigenvalues on the imaginary axis (and in particular is nonsingular). Theorem If x* is hyperbolic and g = f+ p is a small admissible perturbation of f, then g has a unique zero y* near x*. Hyperbolicity

36
Let x*be a hyperbolic equilibrium state for an admissible f. Then the pattern of synchrony ~ x* is rigid if it is the same as ~ y* for the nearby equilibrium y* of any sufficiently small perturbation g. Rigidity More carefully: if x*is a hyperbolic equilibrium state for an admissible f, we can define its rigid pattern of synchrony to be ~ x* rig, where c ~ x* rig d if and only if y c = y d for every nearby equilibrium y* of any sufficiently small perturbation g. Then ~ x* rig is rigid.

37
Rigidity Theorem [Rigid implies balanced] Let x* be a hyperbolic equilibrium state for an admissible f. Then ~ x* rig is balanced. The proof is nontrivial: see Golubitsky, Stewart, and Török. Stronger ResultTransversality Aldis (unpublished) has proved a similar theorem with hyperbolic replaced by transverse, which means that the Jacobian Df| x* has no zero eigenvalues (is nonsingular). This is stronger because Hopf bifurcation points (nonzero imaginary eigenvalues) are permitted.
![Rigidity Theorem [Rigid implies balanced] Let x* be a hyperbolic equilibrium state for an admissible f.](http://images.slideplayer.com/3/799962/slides/slide_37.jpg "Then ~ x* rig is balanced. The proof is nontrivial: see Golubitsky, Stewart, and Török. Stronger ResultTransversality Aldis (unpublished) has proved a similar theorem with hyperbolic replaced by transverse, which means that the Jacobian Df| x* has no zero eigenvalues (is nonsingular). This is stronger because Hopf bifurcation points (nonzero imaginary eigenvalues) are permitted..")
38
Rigid Periodic States Is there an analogous theorem for periodic states? A periodic state x(t) is hyperbolic if its linearized Poincaré map (Floquet map) has no eigenvalues on the unit circle. A pattern of synchrony, or of phase shifts, is rigid if it persists after any sufficiently small admissible perturbation.
 is hyperbolic if its linearized Poincaré map (Floquet map) has no eigenvalues on the unit circle. A pattern of synchrony, or of phase shifts, is rigid if it persists after any sufficiently small admissible perturbation..")
39
Rigid Synchrony Conjecture Conjecture If x(t) is a hyperbolic periodic state, then rigidly synchronous cells have synchronous input cells (up to some input isomorphism). Theorem If the RSC is true, and x(t) is a hyperbolic periodic state, then the relation of rigid synchrony is balanced.
 is a hyperbolic periodic state, then the relation of rigid synchrony is balanced..")
40
Rigid Phase Conjecture Conjecture If x(t) is a hyperbolic periodic state, then rigidly phase-related cells have phase-related input cells (up to some input isomorphism) with the same phase relations. Theorem If the RPC is true, then the quotient network (for the relation of rigid synchrony) has a cyclic group of global symmetries, and the phase relations are among those for a dynamical system with this symmetry.
 has a cyclic group of global symmetries, and the phase relations are among those for a dynamical system with this symmetry..")
41
Dynamics of Networks to be continued... Ian Stewart Mathematics Institute University of Warwick Ian Stewart Mathematics Institute University of Warwick UK-Japan Winter School Dynamics and Complexity

42
Network Each cell has a cell-type and each arrow has an arrow-type, allowing us to require the cells or arrows concerned to have the same structure. In effect these are labels on the cells and arrows. Abstractly they are specified by equivalence relations on the set of cells and the set of arrows.

43
Network Arrows may form loops (same head and tail), and there may be multiple arrows (connecting the same pair of cells). Special case: regular homogeneous networks. These have one type of cell, one type of arrow, and the number of arrows entering each cell is the same. This number is the valency of the network.

44
Regular Homogeneous Network This is a regular homogeneous network of valency 3.

45
Network Enumeration N v=1 v=2 v=3 v=4 v=5 v=6 1111111 23610152128 374418059015823724 4194756915634204122302080827 54768744447221040722682650761843405665412 613012675043242604556967721035590650168613508534834704 Number of topologically distinct regular homogeneous networks on N cells with valency v

46
Network Dynamics To any network we associate a class of admissible vector fields, defining admissible ODEs, which consists of those vector fields F(x) That respect the network structure, and the corresponding ODEs dx/dt = F(x) What does respect the network structure mean?
 That respect the network structure, and the corresponding ODEs dx/dt = F(x) What does respect the network structure mean")
47
Admissible ODEs The input set I(c) of a cell c is the set of all arrows whose head is c. This includes multiple arrows and loops.

48
Admissible ODEs Choose coordinates x c R k for each cell c. (We use R k for simplicity, and because we consider only local bifurcation). Then dx c /dt = f c (x c,x T(I (c)) ) where T(I(c)) is the tuple of tail cells of I(c).
. Then dx c /dt = f c (x c,x T(I (c)) ) where T(I(c)) is the tuple of tail cells of I(c)..")
49
Admissible ODEs dx 1 /dt = f c (x 1,x 1, x 2, x 3, x 3, x 4, x 5, x 5, x 5 ) dx c /dt = f c (x c,x T(I (c)) )
 dx c /dt = f c (x c,x T(I (c)) )")
50
Admissible ODEs for the example network: dx 1 /dt = f(x 1, x 2, x 2, x 3 ) dx 2 /dt = f(x 2, x 3, x 4, x 5 ) dx 3 /dt = f(x 3, x 1, x 3, x 4 ) dx 4 /dt = f(x 4, x 2, x 3, x 5 ) dx 5 /dt = f(x 5, x 2, x 4, x 4 ) Where f satisfies the symmetry condition f(x,u,v,w) is symmetric in u, v, w f(x,u,v,w) is symmetric in u, v, w
 dx 2 /dt = f(x 2, x 3, x 4, x 5 ) dx 3 /dt = f(x 3, x 1, x 3, x 4 ) dx 4 /dt = f(x 4, x 2, x 3, x 5 ) dx 5 /dt = f(x 5, x 2, x 4, x 4 ) Where f satisfies the symmetry condition f(x,u,v,w) is symmetric in u, v, w f(x,u,v,w) is symmetric in u, v, w")
51
dx 1 /dt = f 1 (x 1,x 2,x 4,x 5 ) dx 2 /dt = f 2 (x 2,x 1,x 3,x 5 ) dx 3 /dt = f 3 (x 3,x 1,x 4 ) dx 4 /dt = f 4 (x 4,x 2,x 4 ) dx 5 /dt = f 5 (x 5,x 4 ) domain condition
 dx 2 /dt = f 2 (x 2,x 1,x 3,x 5 ) dx 3 /dt = f 3 (x 3,x 1,x 4 ) dx 4 /dt = f 4 (x 4,x 2,x 4 ) dx 5 /dt = f 5 (x 5,x 4 ) domain condition")
52
dx 1 /dt = f(x 1,x 2,x 4,x 5 ) dx 2 /dt = f(x 2,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 4 ) dx 4 /dt = g(x 4,x 2,x 4 ) dx 5 /dt = h(x 5,x 4 ) pullback condition
 dx 2 /dt = f(x 2,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 4 ) dx 4 /dt = g(x 4,x 2,x 4 ) dx 5 /dt = h(x 5,x 4 ) pullback condition")
53
dx 1 /dt = f(x 1,x 2,x 4,x 5 ) dx 2 /dt = f(x 2,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 4 ) dx 4 /dt = g(x 4,x 2,x 4 ) dx 5 /dt = h(x 5,x 4 ) Vertex group symmetry
 dx 2 /dt = f(x 2,x 1,x 3,x 5 ) dx 3 /dt = g(x 3,x 1,x 4 ) dx 4 /dt = g(x 4,x 2,x 4 ) dx 5 /dt = h(x 5,x 4 ) Vertex group symmetry")
54
[with M.Golubitsky and M.Nicol] Some curious phenomena in coupled cell networks, J. Nonlin. Sci. 14 (2004) 207- 236. [with M.Golubitsky and A.Török] Patterns of synchrony in coupled cell networks with multiple arrows, SIAM J. Appl. Dyn. Sys. 4 (2005) 78-100. [DOI: 10.1137/040612634] [with M.Golubitsky] Nonlinear dynamics of networks: the groupoid formalism, Bull. Amer. Math. Soc. 43 (2006) 305-364. [with M.Parker] Periodic dynamics of coupled cell networks I: rigid patterns of synchrony and phase relations, Dynamical Systems 22 (2007) 389-450. [with M.Parker] Periodic dynamics of coupled cell networks II: cyclic symmetry, Dynamical Systems 23 (2008) 17-41. References
![[with M.Golubitsky and M.Nicol] Some curious phenomena in coupled cell networks, J.](http://images.slideplayer.com/3/799962/slides/slide_54.jpg "Nonlin. Sci. 14 (2004) [with M.Golubitsky and A.Török] Patterns of synchrony in coupled cell networks with multiple arrows, SIAM J. Appl. Dyn. Sys. 4 (2005) [DOI: / ] [with M.Golubitsky] Nonlinear dynamics of networks: the groupoid formalism, Bull. Amer. Math. Soc. 43 (2006) [with M.Parker] Periodic dynamics of coupled cell networks I: rigid patterns of synchrony and phase relations, Dynamical Systems 22 (2007) [with M.Parker] Periodic dynamics of coupled cell networks II: cyclic symmetry, Dynamical Systems 23 (2008) References.")
55
Dynamics of Networks to be continued... Ian Stewart Mathematics Institute University of Warwick Ian Stewart Mathematics Institute University of Warwick UK-Japan Winter School Dynamics and Complexity

Similar presentations