Download presentation
Presentation is loading. Please wait.

1
Transport of Oxygen and carbon dioxide

3
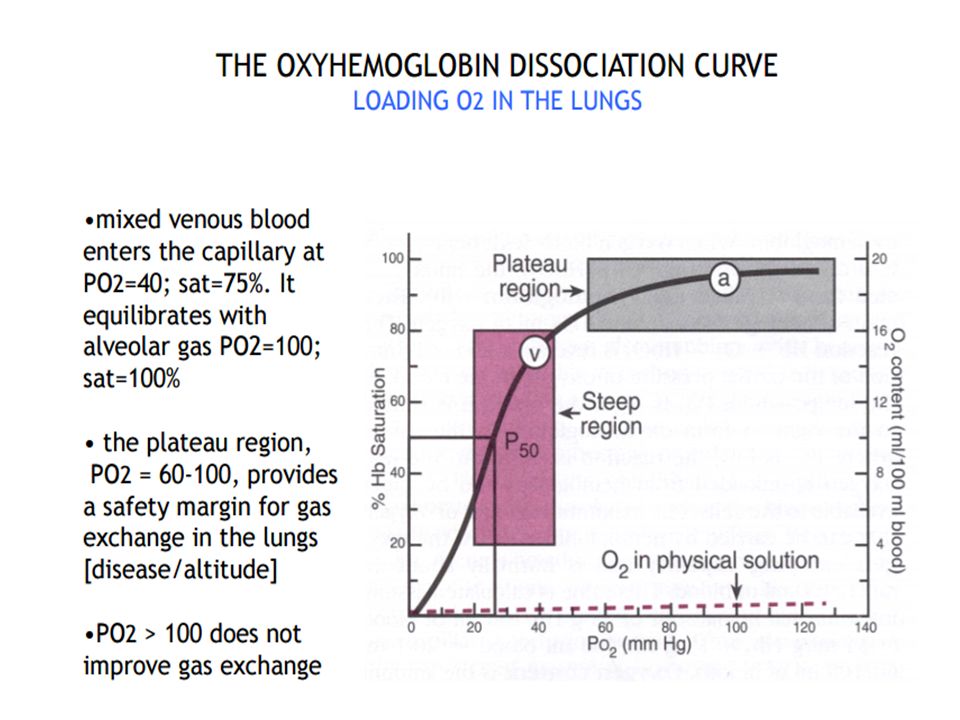
Oxygen uptake in the lungs

5
O2 Transport in blood

6
Dissolved form: Henry's law deals with gases dissolved in solution (e.g., in blood). Both O2 and CO2 are dissolved in blood (a solution) en route to and from the lungs. An important point is that at equilibrium, the partial pressure of a gas in the liquid phase i.e. solution equals the partial pressure in the gas phase. Thus, if alveolar air has a PO2 of 100 mm Hg, then the capillary blood that equilibrates with alveolar air also will have a PO2 of 100 mm Hg.
. Both O2 and CO2 are dissolved in blood (a solution) en route to and from the lungs. An important point is that at equilibrium, the partial pressure of a gas in the liquid phase i.e. solution equals the partial pressure in the gas phase. Thus, if alveolar air has a PO2 of 100 mm Hg, then the capillary blood that equilibrates with alveolar air also will have a PO2 of 100 mm Hg.")
8
The interaction is reversible and is an oxygenation rather than oxidation. DeoxyHb has a relatively low affinity for O2, but each successive O2-binding event produces a conformational change within the protein that incrementally increases the affinity of the other sites

9
Combined form with hemoglobin

11
Molecular basis of the sigmoid shape

13
Diffusion of oxygen from a tissue capillary to the cells
Diffusion of oxygen from a tissue capillary to the cells. (Po2 in interstitial fluid = 40 mm Hg, and in tissue cells = 23 mm Hg.)
")
15
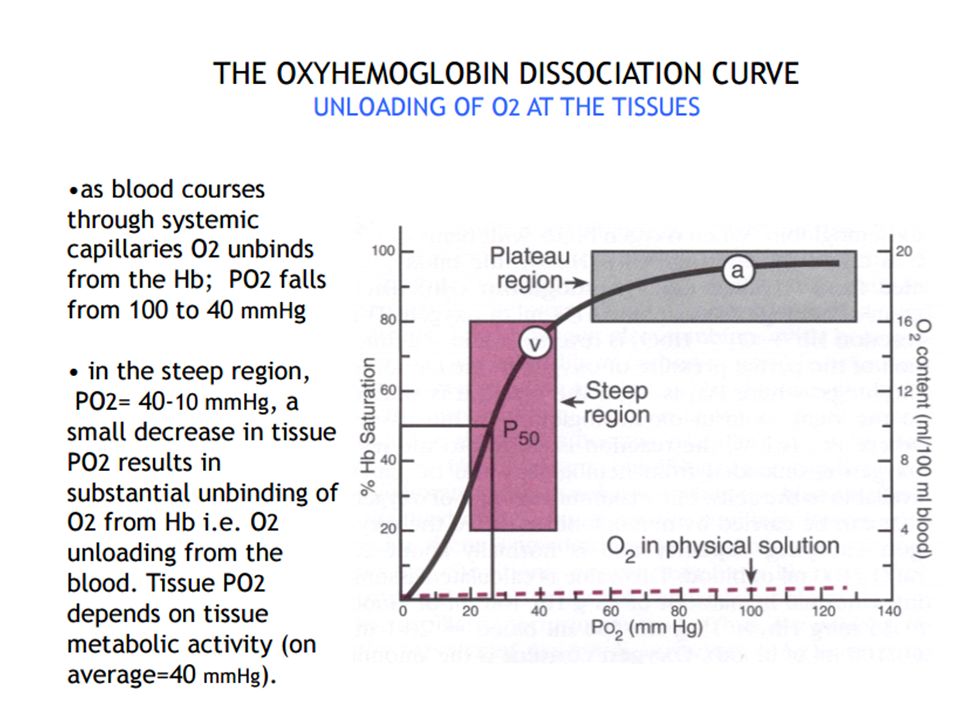
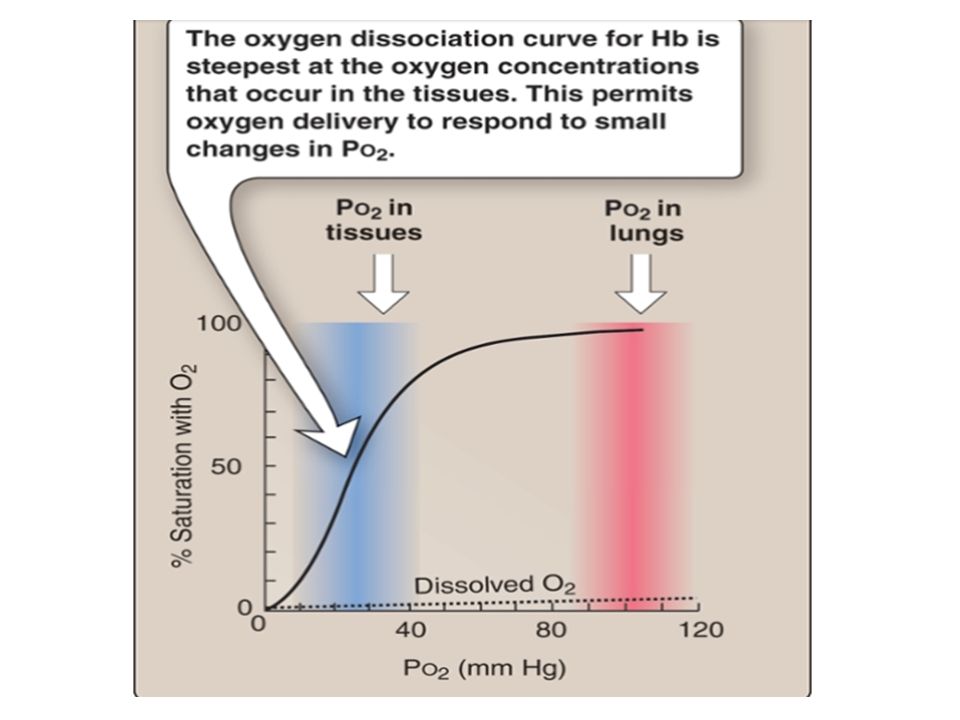
Association: Mixed-venous blood arrives at an alveolus with a Po2 of 40 mm Hg but an O2 saturation of ∼75%. The cooperative nature of O2 binding to Hb means that the single unoccupied heme group has a very high affinity for O2. This allows the site to capture O2 as fast as it can diffuse across the blood–gas interface, simultaneously maintaining a steep pressure gradient for O2 diffusion across the exchange barrier even as equilibration with alveolar gas occurs. Note that the plateau region of the O2 dissociation curve begins at a Po2 of around 60 mm Hg .In practice, this ensures that saturation still occurs if PAo2 is suboptimal (i.e., 60 mm Hg), either because ventilation is impaired or when cardiac output is increased to the point where perfusion becomes limiting. Dissociation: Once blood arrives at a tissue, Hb must release bound O2 and make it available to mitochondria. Transfer is facilitated by the steepness of the pressure gradient between blood and mitochondria, which maintain a local Po2 of ∼3 mm Hg. Hb begins releasing O2 at a Po2 of 60 mm Hg and delivers ∼60% of total as Po2 falls to 20 mm Hg. Each O2 dissociation event lowers the affinity of the remaining heme groups for bound O2, so that if a tissue’s metabolic rate is very high and its need for O2 is increased, unloading occurs with increased efficiency.

17
Hb changes color from dark blue to bright red when O2 binds, which makes it possible to monitor arterial O2-saturation levels using noninvasive pulse oximetry. A light-emitting probe is attached to a finger or ear, then the relative amounts of saturated and desaturated Hb is calculated from the amount of light absorbed at 660 nm and 940 nm, respectively.

18
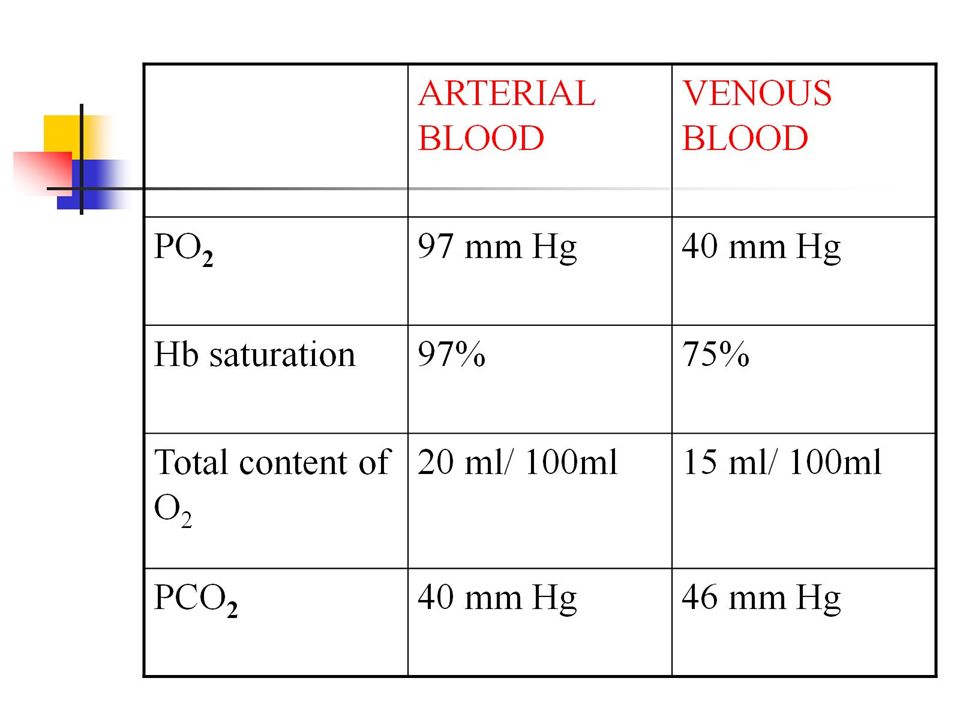
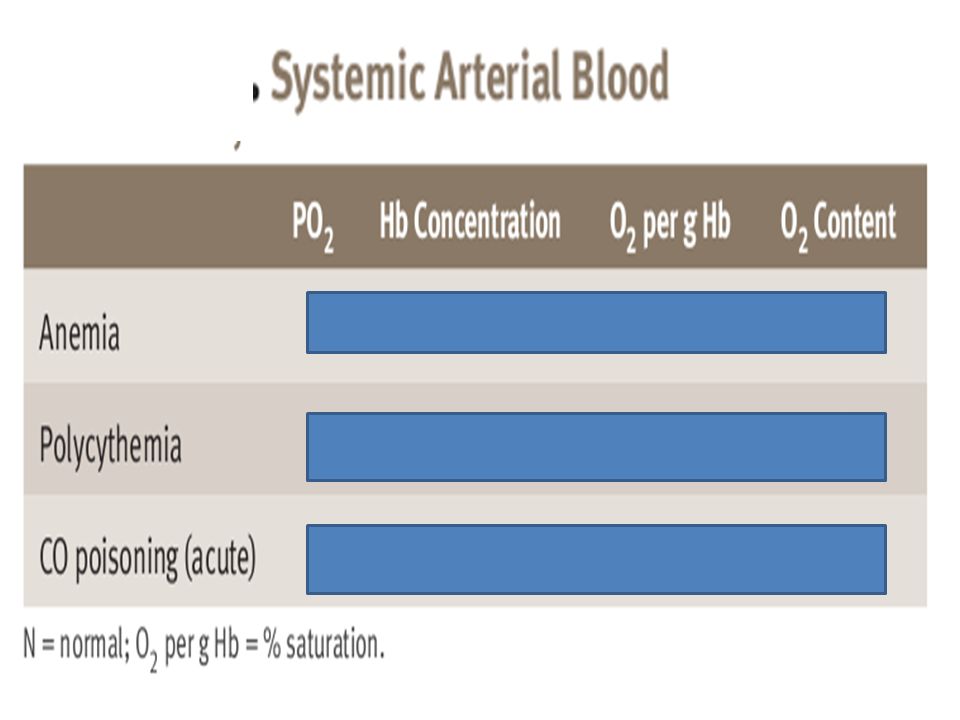
The blood of a normal person contains about 15 grams of hemoglobin in each 100 milliliters of blood, and each gram of hemoglobin can bind with a maximum of 1.34 milliliters of oxygen. There-fore, 15 times 1.34 equals 20.1 On average, the 15 grams of hemoglobin in 100 milliliters of blood can combine with a total of almost exactly 20 milliliters of oxygen if the hemoglobin is 100 per cent saturated On passing through the tissue capillaries, this amount is reduced, on average, to milliliters (Po2of 40 mm Hg, 75 per cent saturated hemoglobin). Thus, under normal conditions, about 5milliliters of oxygen are transported from the lungs to the tissues by each 100 milliliters of blood flow.
. Thus, under normal conditions, about 5milliliters of oxygen are transported from the lungs to the tissues by each 100 milliliters of blood flow.")
19
The blood of a normal person contains about 15 grams of hemoglobin in each 100 milliliters of blood, and each gram of hemoglobin can bind with a maximum of 1.34 milliliters of oxygen. There-fore, 15 times 1.34 equals 20.1 On average, the 15 grams of hemoglobin in 100 milliliters of blood can combine with a total of almost exactly 20 milliliters of oxygen if the hemoglobin is 100 per cent saturated On passing through the tissue capillaries, this amount is reduced, on average, to milliliters (Po2of 40 mm Hg, 75 per cent saturated hemoglobin). Thus, under normal conditions, about 5milliliters of oxygen are transported from the lungs to the tissues by each 100 milliliters of blood flow.
. Thus, under normal conditions, about 5milliliters of oxygen are transported from the lungs to the tissues by each 100 milliliters of blood flow.")
20
P 50

22
Rightward shifts: Metabolism generates heat and CO2 and acidifies the local environment. All three changes reduce Hb’s O2 affinity and cause it to unload O2. The liberated O2 keeps free (dissolved) O2 levels high and maintains a steep pressure gradient between blood and mitochondria even as blood’s O2 stores are being emptied. 2,3-Diphosphoglycerate: 2,3-Diphosphoglycerate (2,3-DPG) is synthesized from 1,3-DPG, which is an intermediate in the glycolytic pathway. 2,3-DPG is abundant in RBCs, its concentration rivaling that of Hb. 2,3-DPG binds preferentially to the deoxygenated form of Hb and stabilizes it, thereby reducing its O2 affinity .The Hb–O2 dissociation curve shifts to the right, and O2 is unloaded. 2,3-DPG and its effects on O2 affinity is a constant in blood, unlike the effects of temperature, CO2, and H+, which typically remain localized to an active tissue.
 is synthesized from 1,3-DPG, which is an intermediate in the glycolytic pathway. 2,3-DPG is abundant in RBCs, its concentration rivaling that of Hb. 2,3-DPG binds preferentially to the deoxygenated form of Hb and stabilizes it, thereby reducing its O2 affinity .The Hb–O2 dissociation curve shifts to the right, and O2 is unloaded. 2,3-DPG and its effects on O2 affinity is a constant in blood, unlike the effects of temperature, CO2, and H+, which typically remain localized to an active tissue.")
23
Chronic hypoxemia caused by pathologic changes in lung function or living at high altitude stimulates 2,3-DPG production. Increased 2,3-DPG levels shift the Hb–O2 dissociation curve even further to the right, which increases the tissue’s accessibility to available O2 .Although 2,3-DPG does reduce the efficiency of O2 loading by Hb in the lungs, the effects are minor and more than offset by the beneficial effects of assisting O2 delivery to tissues If HbA is stripped of 2,3-DPG, its O2-dissociation curve resembles that of HbF. Storing blood causes 2,3-DPG concentrations to decline over the course of a week, causing a leftward shift in the dissociation curve .Although RBCs replenish lost 2,3-DPG within hours to days of transfusion, giving a critically ill patient large volumes of 2,3-DPG–depleted blood presents some difficulties because such blood does not readily give up its O2.

24
HbF’s increased O2 affinity compared with the adult form (HbA) reflects the fact that γ-globins bind 2,3-DPG very weakly. 2,3-DPG normally stabilizes the deoxygenated form of HbA and reduces its affinity. HbF’s inability to bind 2,3-DPG favors O2 loading at low partial pressures.

25
Aerobic metabolism generates CO2 and causes tissue Pco2 to rise
Aerobic metabolism generates CO2 and causes tissue Pco2 to rise. CO2 binds to terminal globin amino groups and decreases Hb’s O2 affinity. The Hb–O2 dissociation curve shifts to the right, and O2 is unloaded. CO2 also dissolves in water to yield free acid, which promotes further O2 unloading via the Bohr effect

26
BOHR EFFECT The PH of the blood falls as its CO2 content increases

27
HbF’s increased O2 affinity compared with the adult form (HbA) reflects the fact that γ-globins bind 2,3-DPG very weakly. 2,3-DPG normally stabilizes the deoxygenated form of HbA and reduces its affinity. HbF’s inability to bind 2,3-DPG favors O2 loading at low partial pressures.

28
CO is formed by combustion of hydrocarbons
CO is formed by combustion of hydrocarbons. Common sources of exposure include automobile exhaust, poorly ventilated heating systems, and smoke. Carboxyhemoglobin comprises up to ∼3% of total Hb in nonsmokers, increasing to 10%–15% in smokers

29
CO poisoning

31
CO poisoning CO decreases O2 bound to hemoglobin and also causes a left shift of the O2-hemoglobin dissociation curve. CO binds to hemoglobin with an affinity that is 250 times that of O2 to form carboxyhemoglobin. the presence of CO decreases the number of O2-binding sites available on hemoglobin. Reduces O2 content of blood and O2 delivery to tissues

35
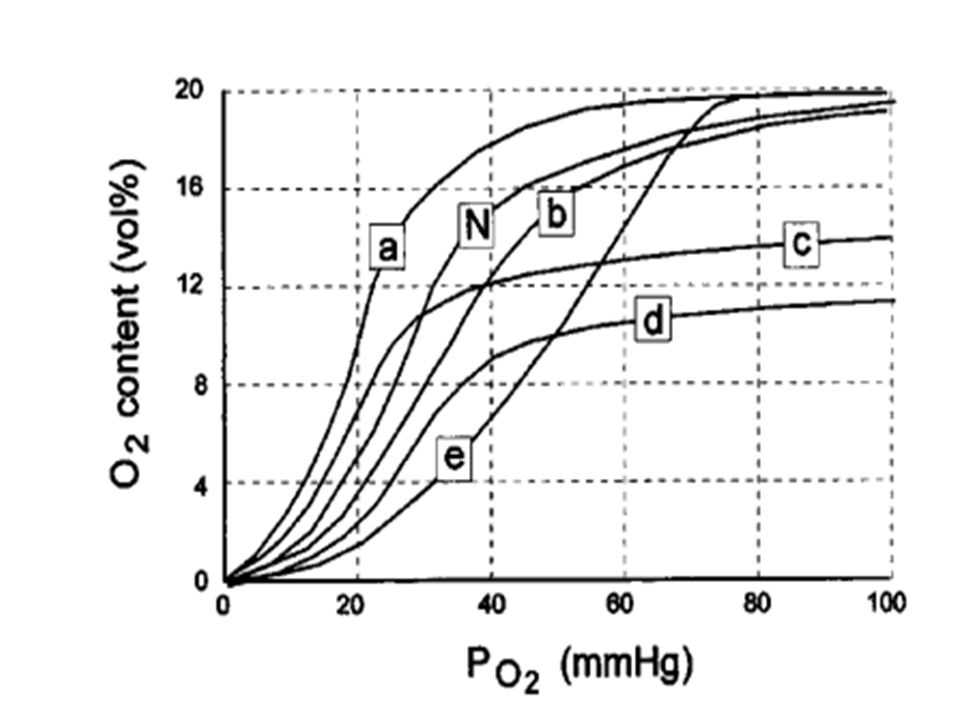
Use the following diagram of oxyhemoglobin saturation curves

36
What is the P50of the oxyhemoglobin curve labeled A in the diagram. a
What is the P50of the oxyhemoglobin curve labeled A in the diagram? a. 80 mmHg b. 60 mmHg c. 40 mmHg d. 30 mmHg e. 20 mmHg Which of the following conditions is most likely to shift the above oxyhemoglobin curve from A to B? a. Increased temperature b. Exercise c. Acclimatization to high altitude d. Hyperventilation e. Metabolic acidosis

37
Which of the following conditions causes a decrease in arterial O2 saturation without a decrease in O2 tension? a. Anemia b. Carbon monoxide poisoning c. A low V/Q ratio d. Hypoventilation e. Right-to-left shunt

39
Which one of the above oxyhemoglobin saturation curves was obtained from fetal blood? a. A b. B c. C d. D e. E

40
Which one of the above oxyhemoglobin saturation curves was obtained from blood exposed to carbon monoxide? a. A b. B c. C d. D e. E

41
TRANSPORT OF CARBON DIOXIDE

43
In the tissues, CO2 is produced from aerobic metabolism
In the tissues, CO2 is produced from aerobic metabolism. CO2 then diffuses across the cell membranes and across the capillary wall, into the red blood cells by simple diffusion, driven by the partial pressure gradient for CO2. Carbonic anhydrase is found in high concentration in red blood cells. It catalyzes the hydration of CO2 to form H2CO3. In the red blood cells, H2CO3 dissociates into H+ and HCO3-. The H+ remains in the red blood cells, where it will be buffered by deoxyhemoglobin, and the HCO3- is transported into the plasma in exchange for Cl- (chloride) i.e. chloride shift. H+ is buffered by deoxyhemoglobin. Deoxyhemoglobin is a better buffer than oxyhemoglobin. The bicarbonate that is formed in the red blood cell is carried in the plasma compartment. All of the reactions described here occur in reverse in the lungs. H+ is released from its buffering sites on deoxyhemoglobin, HCO3 - enters the red blood cells in exchange for Cl-. H+ and HCO3- combine to form H2CO3, and H2CO3 dissociates into CO2 and H2O. The regenerated CO2 and H2O are expired by the lungs
 i.e. chloride shift. H+ is buffered by deoxyhemoglobin. Deoxyhemoglobin is a better buffer than oxyhemoglobin. The bicarbonate that is formed in the red blood cell is carried in the plasma compartment. All of the reactions described here occur in reverse in the lungs. H+ is released from its buffering sites on deoxyhemoglobin, HCO3 - enters the red blood cells in exchange for Cl-. H+ and HCO3- combine to form H2CO3, and H2CO3 dissociates into CO2 and H2O. The regenerated CO2 and H2O are expired by the lungs.")
44
The H+ released during HCO3− formation remains trapped in RBCs by the cell membrane, which is relatively impermeable to cations. This might be expected to lower intracellular pH, but H+ accumulation occurs at the precise moment that Hb is releasing O2 and undergoing a conformational change that favors H+ binding. As noted above (i.e., the Bohr effect), H+ binding actually facilitates O2 unloading by shifting the Hb–O2 dissociation curve to the right and reducing Hb’s affinity for O2. Virtually all of the acid excess caused by loss of HCO3− to the plasma is buffered by Hb. With intracellular H+ kept low by Hb and the Cl−-HCO3− exchanger keeping HCO3− low, the reaction catalyzed by CA remains biased in favor of increased H+ and HCO3− formation. The CO2-carrying capacity of blood increases as a result.
, H+ binding actually facilitates O2 unloading by shifting the Hb–O2 dissociation curve to the right and reducing Hb’s affinity for O2. Virtually all of the acid excess caused by loss of HCO3− to the plasma is buffered by Hb. With intracellular H+ kept low by Hb and the Cl−-HCO3− exchanger keeping HCO3− low, the reaction catalyzed by CA remains biased in favor of increased H+ and HCO3− formation. The CO2-carrying capacity of blood increases as a result..")
45
Transport of Carbon dioxide
Dissolved Carbon Dioxide 7% of the total CO2 content of blood Carbon dioxide is 20 times more soluble in blood than oxygen is. Carbamino Compound Carbon dioxide reacts with terminal amine groups of proteins(Hb) to form carbamino compounds. The protein involved appears to be almost exclusively hemoglobin and this binding is responsible for Bohr effect. Reversly, O2 bound to Hb changes its affinity for CO2, such that when less O2 is bound, the affinity of hemoglobin for CO2 increases called the Haldane effect. About 7% of the total CO2 is carried as carbamino compounds. The attachment sites that bind CO2 are different from the sites that bind O2.
 to form carbamino compounds. The protein involved appears to be almost exclusively hemoglobin and this binding is responsible for Bohr effect. Reversly, O2 bound to Hb changes its affinity for CO2, such that when less O2 is bound, the affinity of hemoglobin for CO2 increases called the Haldane effect. About 7% of the total CO2 is carried as carbamino compounds. The attachment sites that bind CO2 are different from the sites that bind O2.")
46
BOHR EFFECT The PH of the blood falls as its CO2 content increases

47
Bicarbonate About 70% of the CO2 is carried as plasma bicarbonate. In order to convert CO2 into bicarbonate or the reverse, carbonic anhydrase (CA) enzyme must be present. Plasma contains no carbonic anhydrase; therefore, there can be no significant conversion of CO2 to HCO3- in this compartment.
 enzyme must be present. Plasma contains no carbonic anhydrase; therefore, there can be no significant conversion of CO2 to HCO3- in this compartment.")
48
All of the reactions described here occur in reverse in the lungs.
H+ is released from its buffering sites on deoxyhemoglobin, HCO3 - enters the red blood cells in exchange for Cl-. H+ and HCO3- combine to form H2CO3, and H2CO3 dissociates into CO2 and H2O. The regenerated CO2 and H2O are expired by the lungs

50
REVERSED CHLORIDE SHIFT

54
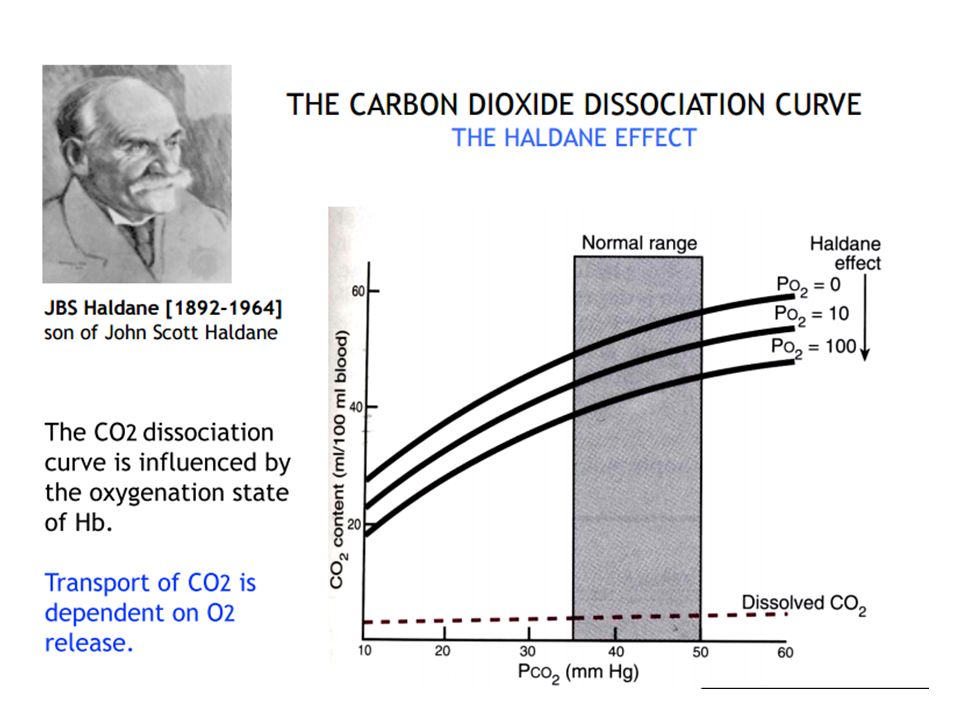
. The degree of Hgb saturation with O2 has a major effect on the CO2 dissociation curve. Although O2 and CO2 bind to Hgb at different sites, deoxygenated Hgb has greater affinity for CO2 than oxygenated Hgb does. Thus, deoxygenated blood (venous blood) freely takes up and transports more CO2 than oxygenated arterial blood does. The deoxygenated Hgb more readily forms carbamino compounds and also more readily binds free H+ ions released during the formation of HCO3-. The effect of changes in oxyhemoglobin saturation on the relationship of CO2 content to Pco2 is referred to as the Haldane effect and is reversed in the lung when O2 is transported from the alveoli to red blood cells. This effect is illustrated by a shift to the left in the CO2 dissociation curve in venous blood as compared with arterial blood
 freely takes up and transports more CO2 than oxygenated arterial blood does. The deoxygenated Hgb more readily forms carbamino compounds and also more readily binds free H+ ions released during the formation of HCO3-. The effect of changes in oxyhemoglobin saturation on the relationship of CO2 content to Pco2 is referred to as the Haldane effect and is reversed in the lung when O2 is transported from the alveoli to red blood cells. This effect is illustrated by a shift to the left in the CO2 dissociation curve in venous blood as compared with arterial blood.")
Similar presentations






















 & CARBONDIOXIDE(CO2)>")



 Chapter 22 Functions of “Blood” Gas Transport Nutrient Transport Excretory Product Transport Cell Signal Transport Hydraulic.>")



