Download presentation
Presentation is loading. Please wait.

1
Metabolism of monosaccharides and disaccharides UNIT II: Intermediary Metabolism

2
Figure 12.1. Galactose and fructose metabolism as part of the essential pathways of energy metabolism.

3
Overview Although many monosaccharides have been identified in nature, only a few sugars appear as metabolic intermediates or as structural components in mammals. Gluc is the most common monosacch consumed by humans, & its metabolism has been discussed However, two other monosacch’s, fructose & galactose, occur in significant amounts in the diet, & make important contributions to energy metabolism In addition, galactose is an important component of cell structural CHO’s

4
II. Fructose metabolism -~ 10% of the calories contained in Western diet are supplied by fructose (~ 50g/day). -The major source of fructose is the disaccharide sucrose, which, when cleaved in intestine, releases equimolar amounts of fructose & glucose -Fructose is also found as a free monosacch in high- fructose corn syrup (55% fructose/45% glucose, which is used to sweeten most cola drinks), in many fruits, & in honey. -Entry of fructose into cells is not insulin-dependent (unlike that of glucose into certain tissues), & in contrast to glucose, fructose does not promote secretion of insulin
. -The major source of fructose is the disaccharide sucrose, which, when cleaved in intestine, releases equimolar amounts of fructose & glucose -Fructose is also found as a free monosacch in high- fructose corn syrup (55% fructose/45% glucose, which is used to sweeten most cola drinks), in many fruits, & in honey. -Entry of fructose into cells is not insulin-dependent (unlike that of glucose into certain tissues), & in contrast to glucose, fructose does not promote secretion of insulin.")
5
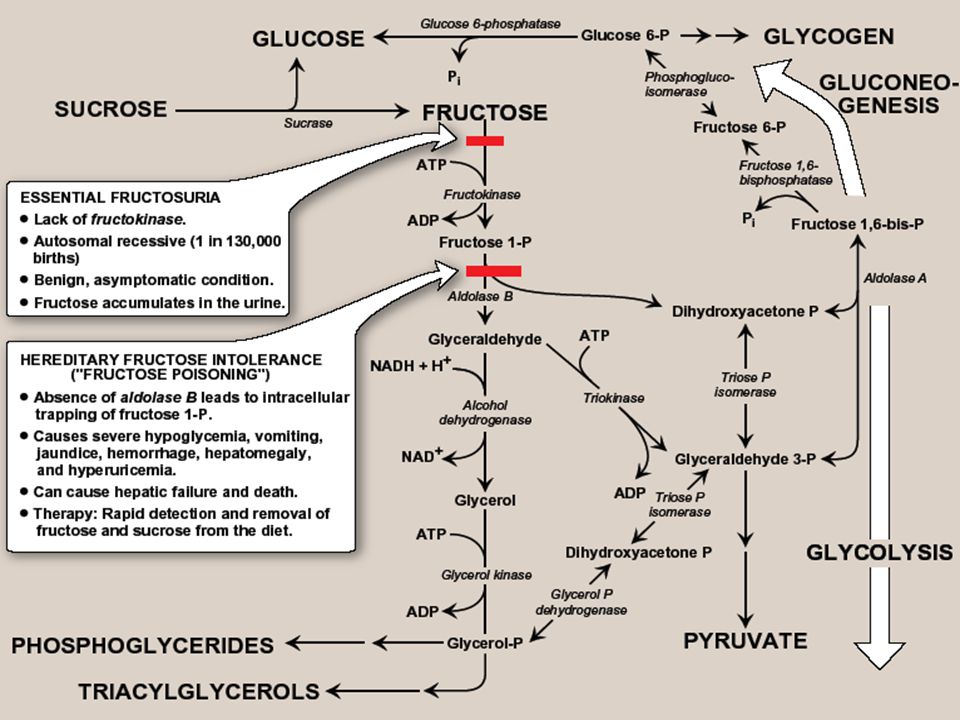
A. Phosphorylation of fructose -For fructose to enter pathways of intermediary metabolism, it must 1 st be phosphorylated. This can be accomplished by either hexokinase or fructokinase (a.k.a ketohexokinase) -Hexokinase phosphorylates gluc in all cells of the body, & several additional hexoses can serve as substrates for this enz. However, it has a low affinity (high Km) for fructose. Therefore, unless intracellular conc. of fructose becomes unusually high, the normal presence of saturating conc’s of gluc means that little fructose is converted to F-6-P by hexokinase -Fructokinase provides the primary mechanism for fructose phospho. It is found in the liver (which processes most of dietary fructose), kidney, & small intestinal mucosa, & converts fructose to F-1-P, using ATP as the P donor Note: these 3 tissues also contain aldolase B
 -Hexokinase phosphorylates gluc in all cells of the body, & several additional hexoses can serve as substrates for this enz. However, it has a low affinity (high Km) for fructose. Therefore, unless intracellular conc. of fructose becomes unusually high, the normal presence of saturating conc’s of gluc means that little fructose is converted to F-6-P by hexokinase -Fructokinase provides the primary mechanism for fructose phospho. It is found in the liver (which processes most of dietary fructose), kidney, & small intestinal mucosa, & converts fructose to F-1-P, using ATP as the P donor Note: these 3 tissues also contain aldolase B.")
6
B. Cleavage of fructose 1-phosphate - F 1-P is not converted to F 1,6-BP as F-6-P, but is cleaved by aldolase B (a.k.a fructose 1- phosphate aldolase) to DHAP & glyceraldehyde Note: both aldolase A (found in all tissues) & aldolase B cleave F 1,6-BP produced during glycolysis to DHAP & GA-3P. - DHAP can directly enter glycolysis or gluconeogenesis, whereas glyceraldehyde can be metabolized by a number of pathways
 to DHAP & glyceraldehyde Note: both aldolase A (found in all tissues) & aldolase B cleave F 1,6-BP produced during glycolysis to DHAP & GA-3P. - DHAP can directly enter glycolysis or gluconeogenesis, whereas glyceraldehyde can be metabolized by a number of pathways.")
7
C. Kinetics of fructose metabolism -The rate of fructose metabolism is more rapid than that of gluc because the trioses formed from F-1-P bypass PFK, the major rate-limiting step in glycolysis Note: loading the liver with fructose, e.g., by intravenous infusion, can significantly elevate the rate of lipogenesis caused by enhanced production of acetyl CoA

8
D. Disorders of fructose metabolism -A deficiency of one of the key enz’s required for the entry of fructose into intermediary metabolic pathways can result in either a benign condition (fructokinase deficiency essential fructosuria), or a severe disturbance of liver & kidney metabolism as a result of aldolase B deficiency (hereditary fructose intolerance, HFI-”fructose poisoning”) which is estimated to occur in 1:20,000 live births -The 1 st symptoms appear when a baby is weaned & begins to be fed food containing sucrose or fructose. F-1-P accumulates, & ATP & Pi levels fall significantly, with adenine being converted to uric acid, causing hyperuricemia
, or a severe disturbance of liver & kidney metabolism as a result of aldolase B deficiency (hereditary fructose intolerance, HFI- fructose poisoning ) which is estimated to occur in 1:20,000 live births -The 1 st symptoms appear when a baby is weaned & begins to be fed food containing sucrose or fructose. F-1-P accumulates, & ATP & Pi levels fall significantly, with adenine being converted to uric acid, causing hyperuricemia.")
9
-The decreased availability of hepatic ATP affects gluconeogenesis (causing hypoglycemia with vomiting), & protein synthesis (causing a decrease in blood clotting factors & other essential proteins) -If fructose (& therefore, sucrose) is not removed from diet, liver failure & death can occur -Diagnosis of HFI can be made on basis of fructose in urine, or by a RFLP test
, & protein synthesis (causing a decrease in blood clotting factors & other essential proteins) -If fructose (& therefore, sucrose) is not removed from diet, liver failure & death can occur -Diagnosis of HFI can be made on basis of fructose in urine, or by a RFLP test")
11
E. Conversion of mannose to fructose 6-phosphate -Mannose, the C-2 epimer of gluc, is an important component of glycoproteins -Hexokinase phosphorylates mannose mannose 6-P, which in turn, is (reversibly) isomerized to F-6-P by phosphomannose isomerase Note: there is little mannose in dietary CHOs. Most intracellular mannose is synthesized from fructose, or is pre-existing mannose produced by degradation of structural CHO’s & salvaged by hexokinase
 isomerized to F-6-P by phosphomannose isomerase Note: there is little mannose in dietary CHOs. Most intracellular mannose is synthesized from fructose, or is pre-existing mannose produced by degradation of structural CHO’s & salvaged by hexokinase.")
12
F. Conversion of glucose to fructose via sorbitol -Most sugars are rapidly phosphorylated following their entry into cells. They are thereby trapped within cells, because organic P’s can’t freely cross memb’s without specific transporters -An alternate mechanism for metabolizing a monosacch is to convert it to a polyol by reduction of an aldehyde group, thereby producing an additional hydoxyl group

13
1. Synthesis of sorbitol: -Aldose reductase reduces glucose, producing sorbitol (glucitol). This enz is found in many tissues, including the lens, retina, Schwann cells of peripheral nerves, liver, kidney, placenta, RBCs, & cells of the ovaries & seminal vesicles. -In cells of liver, ovaries, sperm & seminal vesicles, there is a 2 nd enz, sorbitol dehydrogenase, that can oxidize sorbitol to produce fructose. -The 2-reaction pathway from gluc to fruc in seminal vesicles is for the benefit of sperm cells, which use fruc as a major CHO energy source. -The pathway from sorbitol to fruc in liver provides a mechanism by which any available sorbitol is converted into a substrate that can enter glycolysis or gluconeogenesis

14
Figure 12.4. Sorbitol metabolism

15
2. The effect of hyperglycemia on sorbitol metabolism -Because insulin is not required for entry of gluc into cells listed in previous paragraph, large amounts of gluc may enter these cells during times of hyperglycemia, e.g., in uncontrolled diabetes. -Elevated intracellular gluc conc’s & an adequate supply of NADPH cause aldose reductase to produce a sufficient increase in the amount of sorbitol, which can’t pass efficiently through CMs &, therefore, remains trapped inside cell. -This is exacerbated when sorbitol dehydrogenase is low or absent, e.g., in retina, lens, kidney & nerve cells. As a result, sorbitol accumulates in these cells, causing strong osmotic effects &, therefore, cell swelling as a result of water retention -Some of the pathologic alterations associated with diabetes can be attributed, in part, to this phenomenon, including cataract formation, peripheral neuropathy, & vascular problems leading to nephropathy, & retinopathy

16
III. Galactose metabolism -The major dietary source of galactose is lactose (galactosyl β 1,4-glucose) obtained from milk & milk products. Note: digestion of lactose by β-galactosidase (lactase) of the intestinal mucosal CM was discussed earlier -Some galactose can also be obtained by lysosomal degradation of complex CHOs, such as glycoproteins & glycolipids, which are important memb components. -Like fructose, entry of galactose into cells is not insulin dependent A. Phosphorylation of galactose - Like fruc, galactose must be phosphorylated before it can be further metabolized. Most tissues have a specific enz for this purpose, galactokinase, which produces galactose 1-P. ATP is the P donor.
 obtained from milk & milk products. Note: digestion of lactose by β-galactosidase (lactase) of the intestinal mucosal CM was discussed earlier -Some galactose can also be obtained by lysosomal degradation of complex CHOs, such as glycoproteins & glycolipids, which are important memb components. -Like fructose, entry of galactose into cells is not insulin dependent A. Phosphorylation of galactose - Like fruc, galactose must be phosphorylated before it can be further metabolized. Most tissues have a specific enz for this purpose, galactokinase, which produces galactose 1-P. ATP is the P donor..")
17
B. Formation of UDP-galactose Galactose 1-P can’t enter glycolytic pathway unless it is 1 st converted to UDP-galactose. This occurs as an exchange reaction, in which UMP is removed from UDP- gluc (leaving behind G-1-P), & is then transferred to the galactose 1-P, producing UDP-galactose. The enz which catalyzes this reaction is galactose 1-phosphate uridyl transferase C. Use of UDP-galactose as a carbon source for glycolysis or gluconeogenesis In order for UDP-galactose to enter the mainstream of gluc metabolism, it must 1 st be converted to its C-4 epimer, UDP-gluc, by UDP-hexose 4-epimerase This “new” UDP-gluc (produced from the original UDP- galactose) can then participate in many biosynthetic reactions, as well as being used in uridyl transferase reaction described above, converting another galactose 1-P into UDP-galactose, & releasing G-1-P, whose carbons are those of the original galactose
, & is then transferred to the galactose 1-P, producing UDP-galactose. The enz which catalyzes this reaction is galactose 1-phosphate uridyl transferase C. Use of UDP-galactose as a carbon source for glycolysis or gluconeogenesis In order for UDP-galactose to enter the mainstream of gluc metabolism, it must 1 st be converted to its C-4 epimer, UDP-gluc, by UDP-hexose 4-epimerase This new UDP-gluc (produced from the original UDP- galactose) can then participate in many biosynthetic reactions, as well as being used in uridyl transferase reaction described above, converting another galactose 1-P into UDP-galactose, & releasing G-1-P, whose carbons are those of the original galactose.")
18
Figure 12.6. Structure of UDP-galactose.

19
D. Role of UDP-galactose in biosynthetic reactions -UDP-galactose can serve as the donor of galactose in a number of synthetic pathways, including synthesis of lactose, glycoproteins, glycolipids, & glycosaminoglycans Note: if galactose is not provided by the diet (e.g., when it can’t be released from lactose as a result of a lack of β-galactosidase in people who are lactose-intolerant), all tissue requirements of UDP-galactose can be met by the action of UDP-hexose 4-epimerase on UDP-glucose, which is efficiently produced from G-1-P
, all tissue requirements of UDP-galactose can be met by the action of UDP-hexose 4-epimerase on UDP-glucose, which is efficiently produced from G-1-P.")
20
E. Disorders of galactose metabolism -Galactose 1-phosphate uridyltransferase is missing in individuals with classic galactosemia. In this disorder, galactose 1-P &, therefore, galactose, accumulates in cells. -Physiologic consequences are similar to those found in fructose intolerance, but a broader spectrum of tissues is affected. -The accumulated galactose is shunted into side pathways such as that of galactitol production. This reaction is catalyzed by aldose reductase, the same enz that converts gluc to sorbitol -Note: a more benign form of galactosemia is caused by a deficiency of galactokinase

21
Figure 12.5. Metabolism of galactose.

22
IV. Lactose synthesis -Lactose is a disacch that consists of a molecule of β-galactose attached by β(1→4) linkage to gluc. Therefore, lactose is galactosyl β(1→4)- glucose. -Lactose, known as “milk sugar”, is produced by mammary glands of most mammals. Therefore, milk & other dairy products are the dietary sources of lactose. -Lactose is synthesized in the ER by lactose synthase (UDP-galactose:glucose galactosyltransferase), which transfers galactose from UDP-galactose to gluc, releasing UDP.
 linkage to gluc. Therefore, lactose is galactosyl β(1→4)- glucose. -Lactose, known as milk sugar , is produced by mammary glands of most mammals. Therefore, milk & other dairy products are the dietary sources of lactose. -Lactose is synthesized in the ER by lactose synthase (UDP-galactose:glucose galactosyltransferase), which transfers galactose from UDP-galactose to gluc, releasing UDP..")
23
-This enz is composed of 2 proteins, A & B. protein A is a β-D-galactosyltransferase & is found in a # of body tissues. In tissues other than the lactating mammary gland, this enz transfers galactose from UDP-galactose to N- acetyl-D-glucoasamine, forming the same β(1→4) linkage found in lactose, & producing N- acetyllactosamine, a component of the structurally important N-linked glycoproteins -In contrast, protein B is found only in lactating mammary glands. It is α-lactalbumin, & its synthesis is stimulated by the peptide hormone, prolactin. -Protein B forms a complex with the enz, protein A, changing specificity of that transferase so that lactose, rather than N-acetyllactosamine, is produced
 linkage found in lactose, & producing N- acetyllactosamine, a component of the structurally important N-linked glycoproteins -In contrast, protein B is found only in lactating mammary glands. It is α-lactalbumin, & its synthesis is stimulated by the peptide hormone, prolactin. -Protein B forms a complex with the enz, protein A, changing specificity of that transferase so that lactose, rather than N-acetyllactosamine, is produced.")
24
Figure 12.7. lactose synthesis

25
Summary The major source of fructose is sucrose, which when cleaved releases equimolar amounts of fructose & glucose. Entry of fruc into cells is insulin-independent. Fruc is 1 st phosphorylated to F-1-P by fructokinase, & then cleaved by aldolase B to DHAP & glyceraldehyde. These enz’s are found in liver, kidney, & small intestinal mucosa. A deficiency of fructokinase causes a benign condition (fructosuria), but a deficiency of aldolase B causes hereditary fructose intolerance, in which severe hypoglycemia & liver failure lead to death if the amount of fruc (and therefore, sucrose) in the diet is not severely limited.
, but a deficiency of aldolase B causes hereditary fructose intolerance, in which severe hypoglycemia & liver failure lead to death if the amount of fruc (and therefore, sucrose) in the diet is not severely limited..")
26
Mannose, an important component of glycoproteins, is phosphorylated by hexokinase to mannose-6-P, which is reversibly isomerized to F-6-P by phosphomannose isomerase. Gluc can be reduced to sorbitol (glucitol) by aldose reductase in many tissues, including the lens, retina, Schwann cells, liver, kidney, ovaries, & seminal vesicles. In cells of liver, ovaries, & seminal vesicles, a 2 nd enz, sorbitol dehydrogenase, can oxidize sorbitol to produce fructose. Hyperglycemia results in accumulation of sorbitol in those cells lacking sorbitol dehydrogenase. The resulting osmotic events cause cell swelling, & may contribute to the cataract formation, peripheral neuropathy, nephropathy, & retinopathy seen in diabetes
 by aldose reductase in many tissues, including the lens, retina, Schwann cells, liver, kidney, ovaries, & seminal vesicles. In cells of liver, ovaries, & seminal vesicles, a 2 nd enz, sorbitol dehydrogenase, can oxidize sorbitol to produce fructose. Hyperglycemia results in accumulation of sorbitol in those cells lacking sorbitol dehydrogenase. The resulting osmotic events cause cell swelling, & may contribute to the cataract formation, peripheral neuropathy, nephropathy, & retinopathy seen in diabetes.")
27
The major dietary source of galactose is lactose. The entry of galactose into cells is not insulin-dependent. Galactose is 1 st phosphorylated by galactokinase which produces galactose 1-P. this cpd is converted to UDP- galactose by galactose 1-phosphate uridyltransferase, with the nucleotide supplied by UDP-gluc. A deficiency of this enz causes classic galactosemia. Galactose 1-P accumulates, & excess galactose is converted to galactitol by aldose reductase. This causes liver damage, severe mental retardation, & cataracts. Treatment requires removal of galactose (& therefore, lactose) from the diet. In order for UDP-galactose to enter the mainstream of gluc metabolism, it must be converted to UDP-gluc by UDP-hexose-4-epimerase. This enz can also be used to produce UDP-galactose from UDP-gluc when the former is required for synthesis of structural CHOs
 from the diet. In order for UDP-galactose to enter the mainstream of gluc metabolism, it must be converted to UDP-gluc by UDP-hexose-4-epimerase. This enz can also be used to produce UDP-galactose from UDP-gluc when the former is required for synthesis of structural CHOs.")
28
Lactose is a disacch that consists of galactose & gluc. Milk & other dairy products are the dietary sources of lactose. Lactose is synthesized by lactose synthase from UDP- galactose & gluc in the lactating mammary gland. The enz has two subunits, protein A (which is a galatosyl transferase) found in most cells where it synthesizes N- acetyllactosamine) & protein B (α-lactalbumin, which is found only in the lactating mammary glands, & whose synthesis is stimulated by the peptide hormone, prolactin). When both subunits are present, the transferase produces lactose
 found in most cells where it synthesizes N- acetyllactosamine) & protein B (α-lactalbumin, which is found only in the lactating mammary glands, & whose synthesis is stimulated by the peptide hormone, prolactin). When both subunits are present, the transferase produces lactose.")
29
Figure 12.8. Key concept map for metabolism of fructose and galactose.

Similar presentations


 What is hyperglycemia? Why was there hyperglycemia in this patient?>")







 Lactate or ethanol (anaerobic) (2) Acetyl CoA (aerobic) Acetyl CoA.>")

 polyhydroxy aldehydes. b) polyhydroxy ketones. c) polyhydroxy acids. d) polyhydroxy alcohols.>")






