Download presentation
Presentation is loading. Please wait.

1
Population Structure and Drift Chapter 11

2
Conservation Genetics Illinois Greater Prairie Chicken Illinois Greater Prairie Chicken Tympanuchus cupido pinnatus Tympanuchus cupido pinnatus 200 years ago Illinois was covered with prairie with millions of greater prairie chickens 200 years ago Illinois was covered with prairie with millions of greater prairie chickens Introduction of steel plow decimated prairie habitat Introduction of steel plow decimated prairie habitat By 1994 there were less than 50 greater prairie chickens left By 1994 there were less than 50 greater prairie chickens left Two remnant populations Two remnant populations Illinois Greater Prairie Chicken Illinois Greater Prairie Chicken Tympanuchus cupido pinnatus Tympanuchus cupido pinnatus 200 years ago Illinois was covered with prairie with millions of greater prairie chickens 200 years ago Illinois was covered with prairie with millions of greater prairie chickens Introduction of steel plow decimated prairie habitat Introduction of steel plow decimated prairie habitat By 1994 there were less than 50 greater prairie chickens left By 1994 there were less than 50 greater prairie chickens left Two remnant populations Two remnant populations

5
Conservation Genetics Ban on hunting began in 1933 Ban on hunting began in 1933 In 1960s the two populations were established as sanctuaries In 1960s the two populations were established as sanctuaries In 1970s bird numbers increased In 1970s bird numbers increased By mid-1970s population began to crash again By mid-1970s population began to crash again In 1994 only 6 males were left In 1994 only 6 males were left Why did population decline when habitat was restored? Why did population decline when habitat was restored? Ban on hunting began in 1933 Ban on hunting began in 1933 In 1960s the two populations were established as sanctuaries In 1960s the two populations were established as sanctuaries In 1970s bird numbers increased In 1970s bird numbers increased By mid-1970s population began to crash again By mid-1970s population began to crash again In 1994 only 6 males were left In 1994 only 6 males were left Why did population decline when habitat was restored? Why did population decline when habitat was restored?

7
Random Genetic Drift In finite populations, allele frequencies can fluctuate by chance In finite populations, allele frequencies can fluctuate by chance e.g. populations heavily structured by geography mating not random e.g. populations heavily structured by geography mating not random In finite populations, allele frequencies can fluctuate by chance In finite populations, allele frequencies can fluctuate by chance e.g. populations heavily structured by geography mating not random e.g. populations heavily structured by geography mating not random

8
Consequence Consequence the replacement of old alleles by new ones proceeds in a random fashion not influenced by natural selection the replacement of old alleles by new ones proceeds in a random fashion not influenced by natural selection NONADAPTIVE evolution! NONADAPTIVE evolution! Allele frequencies CAN & DO change from 1 generation to the next Allele frequencies CAN & DO change from 1 generation to the next Consequence Consequence the replacement of old alleles by new ones proceeds in a random fashion not influenced by natural selection the replacement of old alleles by new ones proceeds in a random fashion not influenced by natural selection NONADAPTIVE evolution! NONADAPTIVE evolution! Allele frequencies CAN & DO change from 1 generation to the next Allele frequencies CAN & DO change from 1 generation to the next Random Genetic Drift

9
What are the two MOST important causes of allele substitutions or evolution in populations Random Genetic Drift Random Genetic Drift Natural Selection Natural Selection Random Genetic Drift Random Genetic Drift Natural Selection Natural Selection

10
CoalescenceCoalescence Given enough time, all gene copies in a population are ultimately descended from a single ancestral gene copy Given enough time, all gene copies in a population are ultimately descended from a single ancestral gene copy However, this is also affected by: However, this is also affected by: Mutation Mutation Immigration Immigration Natural Selection Natural Selection Given enough time, all gene copies in a population are ultimately descended from a single ancestral gene copy Given enough time, all gene copies in a population are ultimately descended from a single ancestral gene copy However, this is also affected by: However, this is also affected by: Mutation Mutation Immigration Immigration Natural Selection Natural Selection

11
EXAMPLE OF COALESCENCE

12
CoalescenceCoalescence Implies that the average degree of relationship among individuals increases with the passage of time Implies that the average degree of relationship among individuals increases with the passage of time Thus becomes monomorphic, becomes fixed (freq. of 1.0) Thus becomes monomorphic, becomes fixed (freq. of 1.0) Implies that the average degree of relationship among individuals increases with the passage of time Implies that the average degree of relationship among individuals increases with the passage of time Thus becomes monomorphic, becomes fixed (freq. of 1.0) Thus becomes monomorphic, becomes fixed (freq. of 1.0)
 Thus becomes monomorphic, becomes fixed (freq. of 1.0) Implies that the average degree of relationship among individuals increases with the passage of time Implies that the average degree of relationship among individuals increases with the passage of time Thus becomes monomorphic, becomes fixed (freq. of 1.0) Thus becomes monomorphic, becomes fixed (freq. of 1.0).")
13
CoalescenceCoalescence If we have alleles p and q where p=0.5 and q=0.5 If we have alleles p and q where p=0.5 and q=0.5 What would be the probability that allele q is passed on to the next generation? What would be the probability that allele q is passed on to the next generation? If we have alleles p and q where p=0.9 and q=0.1 If we have alleles p and q where p=0.9 and q=0.1 A population will eventually become monomorphic and this equals the initial frequency of that allele A population will eventually become monomorphic and this equals the initial frequency of that allele If we have alleles p and q where p=0.5 and q=0.5 If we have alleles p and q where p=0.5 and q=0.5 What would be the probability that allele q is passed on to the next generation? What would be the probability that allele q is passed on to the next generation? If we have alleles p and q where p=0.9 and q=0.1 If we have alleles p and q where p=0.9 and q=0.1 A population will eventually become monomorphic and this equals the initial frequency of that allele A population will eventually become monomorphic and this equals the initial frequency of that allele

14
CoalescenceCoalescence How long does it take? How long does it take? The mean time back to common ancestry of all gene copies in the population is 2N generations The mean time back to common ancestry of all gene copies in the population is 2N generations Consider the probability of q being passed to the next generation when there was a small number of mating individuals....say 10...??? Consider the probability of q being passed to the next generation when there was a small number of mating individuals....say 10...??? How long does it take? How long does it take? The mean time back to common ancestry of all gene copies in the population is 2N generations The mean time back to common ancestry of all gene copies in the population is 2N generations Consider the probability of q being passed to the next generation when there was a small number of mating individuals....say 10...??? Consider the probability of q being passed to the next generation when there was a small number of mating individuals....say 10...???

15
Genetic Drift 1.Alleles fluctuate at random, and eventually become fixed 2.Genetic variation is lost 3.Initially similar populations diverge in allele frequency, and may become fixed for different alleles (all individuals homozygous) 4.The probability, at time t, that an allele will eventually become fixed equals the frequency of the allele at that time 5.The rate at which these events occur is greater, the smaller the population 1.Alleles fluctuate at random, and eventually become fixed 2.Genetic variation is lost 3.Initially similar populations diverge in allele frequency, and may become fixed for different alleles (all individuals homozygous) 4.The probability, at time t, that an allele will eventually become fixed equals the frequency of the allele at that time 5.The rate at which these events occur is greater, the smaller the population
 4.The probability, at time t, that an allele will eventually become fixed equals the frequency of the allele at that time 5.The rate at which these events occur is greater, the smaller the population 1.Alleles fluctuate at random, and eventually become fixed 2.Genetic variation is lost 3.Initially similar populations diverge in allele frequency, and may become fixed for different alleles (all individuals homozygous) 4.The probability, at time t, that an allele will eventually become fixed equals the frequency of the allele at that time 5.The rate at which these events occur is greater, the smaller the population")
16
N = 4 N = 40 N = 400

17
Drift Simulator http://darwin.eeb.uconn.edu/simulati ons/drift.html http://darwin.eeb.uconn.edu/simulati ons/drift.html

18
Evolution by Genetic Drift In an adult population there are 2N gene copies In an adult population there are 2N gene copies The variance in allele frequencies: The variance in allele frequencies: V = p (1-p) / 2N (where 2 refers to the diploid condition and N refers to the size of the population) What is this telling us? In an adult population there are 2N gene copies In an adult population there are 2N gene copies The variance in allele frequencies: The variance in allele frequencies: V = p (1-p) / 2N (where 2 refers to the diploid condition and N refers to the size of the population) What is this telling us?
 / 2N (where 2 refers to the diploid condition and N refers to the size of the population) What is this telling us .")
19
Evolution by Genetic Drift is FASTER in small populations The smaller the N (pop. size) the greater the genetic variance from generation to generation! The smaller the N (pop. size) the greater the genetic variance from generation to generation! The probability, at time t, that an allele will ultimately become fixed equals its frequency at that time. The probability, at time t, that an allele will ultimately become fixed equals its frequency at that time. If a new mutation arises in a population, the probability it will be fixed in that population at time t is: If a new mutation arises in a population, the probability it will be fixed in that population at time t is: p t = ½ N p t = ½ N this is the likelihood of reaching p=1 (fixation) Clearly, if the population is small, fixation will occur faster Clearly, if the population is small, fixation will occur faster The smaller the N (pop. size) the greater the genetic variance from generation to generation! The smaller the N (pop. size) the greater the genetic variance from generation to generation! The probability, at time t, that an allele will ultimately become fixed equals its frequency at that time. The probability, at time t, that an allele will ultimately become fixed equals its frequency at that time. If a new mutation arises in a population, the probability it will be fixed in that population at time t is: If a new mutation arises in a population, the probability it will be fixed in that population at time t is: p t = ½ N p t = ½ N this is the likelihood of reaching p=1 (fixation) Clearly, if the population is small, fixation will occur faster Clearly, if the population is small, fixation will occur faster
 the greater the genetic variance from generation to generation. The smaller the N (pop. size) the greater the genetic variance from generation to generation. The probability, at time t, that an allele will ultimately become fixed equals its frequency at that time. The probability, at time t, that an allele will ultimately become fixed equals its frequency at that time. If a new mutation arises in a population, the probability it will be fixed in that population at time t is: If a new mutation arises in a population, the probability it will be fixed in that population at time t is: p t = ½ N p t = ½ N this is the likelihood of reaching p=1 (fixation) Clearly, if the population is small, fixation will occur faster Clearly, if the population is small, fixation will occur faster The smaller the N (pop. size) the greater the genetic variance from generation to generation. The smaller the N (pop. size) the greater the genetic variance from generation to generation. The probability, at time t, that an allele will ultimately become fixed equals its frequency at that time. The probability, at time t, that an allele will ultimately become fixed equals its frequency at that time. If a new mutation arises in a population, the probability it will be fixed in that population at time t is: If a new mutation arises in a population, the probability it will be fixed in that population at time t is: p t = ½ N p t = ½ N this is the likelihood of reaching p=1 (fixation) Clearly, if the population is small, fixation will occur faster Clearly, if the population is small, fixation will occur faster.")
20
Evolution by Drift Random walk of the drunk Random walk of the drunk The probability, at time t, that an allele will ultimately be fixed equals it frequency (p t ) at that time. The probability, at time t, that an allele will ultimately be fixed equals it frequency (p t ) at that time. Random walk of the drunk Random walk of the drunk The probability, at time t, that an allele will ultimately be fixed equals it frequency (p t ) at that time. The probability, at time t, that an allele will ultimately be fixed equals it frequency (p t ) at that time.
 at that time. Random walk of the drunk Random walk of the drunk The probability, at time t, that an allele will ultimately be fixed equals it frequency (p t ) at that time. The probability, at time t, that an allele will ultimately be fixed equals it frequency (p t ) at that time..")
21
Structured Populations The Concept of Demes and metapopulations The Concept of Demes and metapopulations

22
Structured Populations

23
Demes and metapopulations Demes and metapopulations the equation to predict the probability of a new mutation becoming fixed is still ½ N the equation to predict the probability of a new mutation becoming fixed is still ½ N if k=# of populations if k=# of populations 2Nk=total # gene copies in population 2Nk=total # gene copies in population Overall number of populations fixed for p: Overall number of populations fixed for p: pk(2N) [where 2N = #genes per population) pk(2N) [where 2N = #genes per population) Demes and metapopulations Demes and metapopulations the equation to predict the probability of a new mutation becoming fixed is still ½ N the equation to predict the probability of a new mutation becoming fixed is still ½ N if k=# of populations if k=# of populations 2Nk=total # gene copies in population 2Nk=total # gene copies in population Overall number of populations fixed for p: Overall number of populations fixed for p: pk(2N) [where 2N = #genes per population) pk(2N) [where 2N = #genes per population)
 [where 2N = #genes per population) pk(2N) [where 2N = #genes per population) Demes and metapopulations Demes and metapopulations the equation to predict the probability of a new mutation becoming fixed is still ½ N the equation to predict the probability of a new mutation becoming fixed is still ½ N if k=# of populations if k=# of populations 2Nk=total # gene copies in population 2Nk=total # gene copies in population Overall number of populations fixed for p: Overall number of populations fixed for p: pk(2N) [where 2N = #genes per population) pk(2N) [where 2N = #genes per population)")
24
frequency of, say the allele p, is summed over all metapopulations p2Nk / 2Nk frequency of, say the allele p, is summed over all metapopulations p2Nk / 2Nk This equals p, the initial frequency! This equals p, the initial frequency! frequency of, say the allele p, is summed over all metapopulations p2Nk / 2Nk frequency of, say the allele p, is summed over all metapopulations p2Nk / 2Nk This equals p, the initial frequency! This equals p, the initial frequency! Structured Populations

25
Thus one consequence of drift is the smaller the population size, the shorter the average time to fixation Thus one consequence of drift is the smaller the population size, the shorter the average time to fixation For diploid populations, a newly arisen neutral allele mutation takes (on average) 4N generations to become fixed For diploid populations, a newly arisen neutral allele mutation takes (on average) 4N generations to become fixed Thus one consequence of drift is the smaller the population size, the shorter the average time to fixation Thus one consequence of drift is the smaller the population size, the shorter the average time to fixation For diploid populations, a newly arisen neutral allele mutation takes (on average) 4N generations to become fixed For diploid populations, a newly arisen neutral allele mutation takes (on average) 4N generations to become fixed Structured Populations
 4N generations to become fixed For diploid populations, a newly arisen neutral allele mutation takes (on average) 4N generations to become fixed Thus one consequence of drift is the smaller the population size, the shorter the average time to fixation Thus one consequence of drift is the smaller the population size, the shorter the average time to fixation For diploid populations, a newly arisen neutral allele mutation takes (on average) 4N generations to become fixed For diploid populations, a newly arisen neutral allele mutation takes (on average) 4N generations to become fixed Structured Populations")
26
HeterozygosityHeterozygosity Heterozygosity is maximal when all alleles are equal (p=0.5 and q=0.5) Heterozygosity is maximal when all alleles are equal (p=0.5 and q=0.5) Alleles may drift from low frequency to high frequency or even fixation in which case: Alleles may drift from low frequency to high frequency or even fixation in which case: Heterozygosity initially increases (as the frequencies start to approach 50% p and q) then starts to decrease as one of the two alleles drift toward fixation Heterozygosity initially increases (as the frequencies start to approach 50% p and q) then starts to decrease as one of the two alleles drift toward fixation In a metapopulation, different alleles become fixed in different demes (or metapopulations) each of which declines in Heterozygosity In a metapopulation, different alleles become fixed in different demes (or metapopulations) each of which declines in Heterozygosity Alleles may drift from low frequency to high frequency or even fixation Alleles may drift from low frequency to high frequency or even fixation Heterozygosity is maximal when all alleles are equal (p=0.5 and q=0.5) Heterozygosity is maximal when all alleles are equal (p=0.5 and q=0.5) Alleles may drift from low frequency to high frequency or even fixation in which case: Alleles may drift from low frequency to high frequency or even fixation in which case: Heterozygosity initially increases (as the frequencies start to approach 50% p and q) then starts to decrease as one of the two alleles drift toward fixation Heterozygosity initially increases (as the frequencies start to approach 50% p and q) then starts to decrease as one of the two alleles drift toward fixation In a metapopulation, different alleles become fixed in different demes (or metapopulations) each of which declines in Heterozygosity In a metapopulation, different alleles become fixed in different demes (or metapopulations) each of which declines in Heterozygosity Alleles may drift from low frequency to high frequency or even fixation Alleles may drift from low frequency to high frequency or even fixation
 Heterozygosity is maximal when all alleles are equal (p=0.5 and q=0.5) Alleles may drift from low frequency to high frequency or even fixation in which case: Alleles may drift from low frequency to high frequency or even fixation in which case: Heterozygosity initially increases (as the frequencies start to approach 50% p and q) then starts to decrease as one of the two alleles drift toward fixation Heterozygosity initially increases (as the frequencies start to approach 50% p and q) then starts to decrease as one of the two alleles drift toward fixation In a metapopulation, different alleles become fixed in different demes (or metapopulations) each of which declines in Heterozygosity In a metapopulation, different alleles become fixed in different demes (or metapopulations) each of which declines in Heterozygosity Alleles may drift from low frequency to high frequency or even fixation Alleles may drift from low frequency to high frequency or even fixation Heterozygosity is maximal when all alleles are equal (p=0.5 and q=0.5) Heterozygosity is maximal when all alleles are equal (p=0.5 and q=0.5) Alleles may drift from low frequency to high frequency or even fixation in which case: Alleles may drift from low frequency to high frequency or even fixation in which case: Heterozygosity initially increases (as the frequencies start to approach 50% p and q) then starts to decrease as one of the two alleles drift toward fixation Heterozygosity initially increases (as the frequencies start to approach 50% p and q) then starts to decrease as one of the two alleles drift toward fixation In a metapopulation, different alleles become fixed in different demes (or metapopulations) each of which declines in Heterozygosity In a metapopulation, different alleles become fixed in different demes (or metapopulations) each of which declines in Heterozygosity Alleles may drift from low frequency to high frequency or even fixation Alleles may drift from low frequency to high frequency or even fixation")
27
During this process in a metapopulation or deme system of structured populations, heterozygosity and homozygosity retain apparent H-W equilibrium genotype frequencies within each population BUT the metapopulation suffers a net DECREASE in Heterozygosity! During this process in a metapopulation or deme system of structured populations, heterozygosity and homozygosity retain apparent H-W equilibrium genotype frequencies within each population BUT the metapopulation suffers a net DECREASE in Heterozygosity! Heterozygosity and Drift

28
What is Predicted based on Genetic Drift Simulations... p = 0.5 q = 0.5 N = 16 2N = 32 Estimates fixation by 19 generations! p = 0.5 q = 0.5 N = 16 2N = 32 Estimates fixation by 19 generations!

29
The Real Experimental Data

30
Effective population size N e The reality of population ecology and demography can affect the reality of chance mating within populations, decreasing the effective population size The reality of population ecology and demography can affect the reality of chance mating within populations, decreasing the effective population size 1.Variation in number of progeny 2.skewed sex ratios 3.overlapping generations 4.fluctuations in population size The reality of population ecology and demography can affect the reality of chance mating within populations, decreasing the effective population size The reality of population ecology and demography can affect the reality of chance mating within populations, decreasing the effective population size 1.Variation in number of progeny 2.skewed sex ratios 3.overlapping generations 4.fluctuations in population size

31
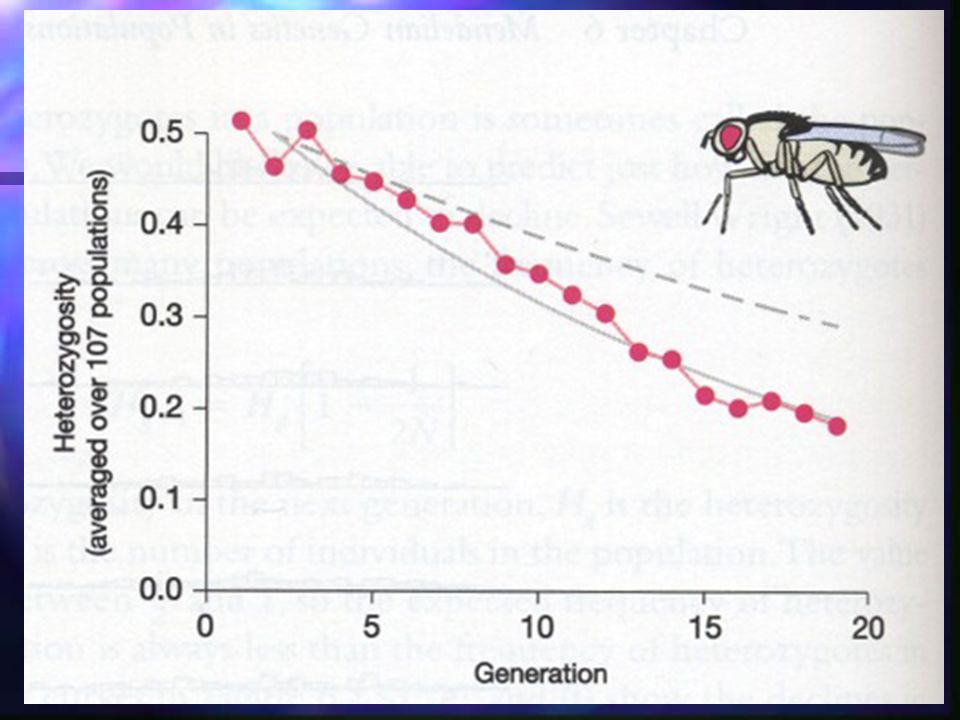
Genetic Drift Experimental study on Drosophila Average heterozygosity fell over time Decrease in heterozygosity did not match predictions exactly Fell at rate predicted for population size of 9, not 16!!! Effective population size was only 9 Some individuals could have died Some males may have been rejected by females Theory of genetic drift can make testable predictions about behavior of alleles in finite populations Experimental study on Drosophila Average heterozygosity fell over time Decrease in heterozygosity did not match predictions exactly Fell at rate predicted for population size of 9, not 16!!! Effective population size was only 9 Some individuals could have died Some males may have been rejected by females Theory of genetic drift can make testable predictions about behavior of alleles in finite populations

33
Founder Effects Genetic bottlenecks Genetic bottlenecks population going through a population crash, followed by a population increase This has massive consequences on the size of the gene pool after population increase, and thus, drastically affects N e Genetic bottlenecks Genetic bottlenecks population going through a population crash, followed by a population increase This has massive consequences on the size of the gene pool after population increase, and thus, drastically affects N e

34
Founder Effects N = number of founders r = pop. increase WE SEE: if “r” is small, Heterozygosity declines faster! N = number of founders r = pop. increase WE SEE: if “r” is small, Heterozygosity declines faster!

35
InbreedingInbreeding Autozygous Autozygous homozygous only and identical by descent homozygous only and identical by descent Allozygous Allozygous either homozygous or heterozygous but not identical by descent either homozygous or heterozygous but not identical by descent An inbred population is one in which the probability that an individual is autozygous is greater, as a consequence of mating among relatives, than in a panmictic population An inbred population is one in which the probability that an individual is autozygous is greater, as a consequence of mating among relatives, than in a panmictic population Autozygous Autozygous homozygous only and identical by descent homozygous only and identical by descent Allozygous Allozygous either homozygous or heterozygous but not identical by descent either homozygous or heterozygous but not identical by descent An inbred population is one in which the probability that an individual is autozygous is greater, as a consequence of mating among relatives, than in a panmictic population An inbred population is one in which the probability that an individual is autozygous is greater, as a consequence of mating among relatives, than in a panmictic population

36
InbreedingInbreeding Inbreeding coeffecient = F the probability that an individual taken at random from a population that is autozygous F is the fraction that is autozygous and 1-F is the fraction that is allozygous F is the fraction that is autozygous and 1-F is the fraction that is allozygous Inbreeding coeffecient = F the probability that an individual taken at random from a population that is autozygous F is the fraction that is autozygous and 1-F is the fraction that is allozygous F is the fraction that is autozygous and 1-F is the fraction that is allozygous

37
InbreedingInbreeding With two alleles A 1 and A 2 with frequencies p and q the probability that an individual is allozygous and A 1 A 1 = (1-F)p 2 With two alleles A 1 and A 2 with frequencies p and q the probability that an individual is allozygous and A 1 A 1 = (1-F)p 2 A 1 A 2 = (1-F)2pq the fraction of heterozygotes (although by definition, they are allozygous) A 1 A 2 = (1-F)2pq the fraction of heterozygotes (although by definition, they are allozygous) A 2 A 2 = (1-F)q 2 A 2 A 2 = (1-F)q 2 Thus the fraction that is autozygous and A 1 A 1 = F(p) and thus, A 2 A 2 = F(q) Thus the fraction that is autozygous and A 1 A 1 = F(p) and thus, A 2 A 2 = F(q) With two alleles A 1 and A 2 with frequencies p and q the probability that an individual is allozygous and A 1 A 1 = (1-F)p 2 With two alleles A 1 and A 2 with frequencies p and q the probability that an individual is allozygous and A 1 A 1 = (1-F)p 2 A 1 A 2 = (1-F)2pq the fraction of heterozygotes (although by definition, they are allozygous) A 1 A 2 = (1-F)2pq the fraction of heterozygotes (although by definition, they are allozygous) A 2 A 2 = (1-F)q 2 A 2 A 2 = (1-F)q 2 Thus the fraction that is autozygous and A 1 A 1 = F(p) and thus, A 2 A 2 = F(q) Thus the fraction that is autozygous and A 1 A 1 = F(p) and thus, A 2 A 2 = F(q)
p 2 With two alleles A 1 and A 2 with frequencies p and q the probability that an individual is allozygous and A 1 A 1 = (1-F)p 2 A 1 A 2 = (1-F)2pq the fraction of heterozygotes (although by definition, they are allozygous) A 1 A 2 = (1-F)2pq the fraction of heterozygotes (although by definition, they are allozygous) A 2 A 2 = (1-F)q 2 A 2 A 2 = (1-F)q 2 Thus the fraction that is autozygous and A 1 A 1 = F(p) and thus, A 2 A 2 = F(q) Thus the fraction that is autozygous and A 1 A 1 = F(p) and thus, A 2 A 2 = F(q) With two alleles A 1 and A 2 with frequencies p and q the probability that an individual is allozygous and A 1 A 1 = (1-F)p 2 With two alleles A 1 and A 2 with frequencies p and q the probability that an individual is allozygous and A 1 A 1 = (1-F)p 2 A 1 A 2 = (1-F)2pq the fraction of heterozygotes (although by definition, they are allozygous) A 1 A 2 = (1-F)2pq the fraction of heterozygotes (although by definition, they are allozygous) A 2 A 2 = (1-F)q 2 A 2 A 2 = (1-F)q 2 Thus the fraction that is autozygous and A 1 A 1 = F(p) and thus, A 2 A 2 = F(q) Thus the fraction that is autozygous and A 1 A 1 = F(p) and thus, A 2 A 2 = F(q)")
38
InbreedingInbreeding Notice, the frequency of homozygotes is higher! Notice, the frequency of homozygotes is higher! F increases over generations F increases over generations Frequency of Heterozygosity is ½ in each successive generation Frequency of Heterozygosity is ½ in each successive generation Notice, the frequency of homozygotes is higher! Notice, the frequency of homozygotes is higher! F increases over generations F increases over generations Frequency of Heterozygosity is ½ in each successive generation Frequency of Heterozygosity is ½ in each successive generation

39
Genetic Consequences of Inbreeding 1.Inbreeding redistributes the alleles to the homozygous condition genotype freqs. change but allele frequencies do not change 2.Genetic variance of a phenotypic character within a population is usually increased 3.inbreeding depression increase in homozygous recessive alleles (likely to increase deleterious phenotypes) 4.Inbreeding promotes linkage disequilibrium nonrandom association of alleles at different loci 1.Inbreeding redistributes the alleles to the homozygous condition genotype freqs. change but allele frequencies do not change 2.Genetic variance of a phenotypic character within a population is usually increased 3.inbreeding depression increase in homozygous recessive alleles (likely to increase deleterious phenotypes) 4.Inbreeding promotes linkage disequilibrium nonrandom association of alleles at different loci
 4.Inbreeding promotes linkage disequilibrium nonrandom association of alleles at different loci 1.Inbreeding redistributes the alleles to the homozygous condition genotype freqs. change but allele frequencies do not change 2.Genetic variance of a phenotypic character within a population is usually increased 3.inbreeding depression increase in homozygous recessive alleles (likely to increase deleterious phenotypes) 4.Inbreeding promotes linkage disequilibrium nonrandom association of alleles at different loci.")
40
Genetic Drift VS Inbreeding Genetic Drift Genetic Drift Allele frequencies change Allele frequencies change Inbreeding Inbreeding Allele frequencies do not change but genotype frequencies (homozygous vs. heterozygotes) DO change! Allele frequencies do not change but genotype frequencies (homozygous vs. heterozygotes) DO change! Genetic Drift Genetic Drift Allele frequencies change Allele frequencies change Inbreeding Inbreeding Allele frequencies do not change but genotype frequencies (homozygous vs. heterozygotes) DO change! Allele frequencies do not change but genotype frequencies (homozygous vs. heterozygotes) DO change!
 DO change. Allele frequencies do not change but genotype frequencies (homozygous vs. heterozygotes) DO change. Genetic Drift Genetic Drift Allele frequencies change Allele frequencies change Inbreeding Inbreeding Allele frequencies do not change but genotype frequencies (homozygous vs. heterozygotes) DO change. Allele frequencies do not change but genotype frequencies (homozygous vs. heterozygotes) DO change!.")
41
Gene flow Natural populations of a species typically are not completely isolated, but instead exchange genes with one another to a greater or lesser extent Natural populations of a species typically are not completely isolated, but instead exchange genes with one another to a greater or lesser extent Gene flow, if unopposed by other factors, homogenizes a population Gene flow, if unopposed by other factors, homogenizes a population Natural populations of a species typically are not completely isolated, but instead exchange genes with one another to a greater or lesser extent Natural populations of a species typically are not completely isolated, but instead exchange genes with one another to a greater or lesser extent Gene flow, if unopposed by other factors, homogenizes a population Gene flow, if unopposed by other factors, homogenizes a population

42
Models of Gene flow Island models Island models Stepping stone models Stepping stone models Isolation by distance models Isolation by distance models Island models Island models Stepping stone models Stepping stone models Isolation by distance models Isolation by distance models

43
Gene flow Homogenizes the populations within a species (unopposed by other forces) Homogenizes the populations within a species (unopposed by other forces) Rate of gene flow = m Rate of gene flow = m A 1 varies among populations (p i ) resident A 1 varies among populations (p i ) resident p i and the average allele frequency is p (source) p i and the average allele frequency is p (source) within population i (p i ), a proportion of m of the gene copies enter from other populations, and the frequency is p within population i (p i ), a proportion of m of the gene copies enter from other populations, and the frequency is p Homogenizes the populations within a species (unopposed by other forces) Homogenizes the populations within a species (unopposed by other forces) Rate of gene flow = m Rate of gene flow = m A 1 varies among populations (p i ) resident A 1 varies among populations (p i ) resident p i and the average allele frequency is p (source) p i and the average allele frequency is p (source) within population i (p i ), a proportion of m of the gene copies enter from other populations, and the frequency is p within population i (p i ), a proportion of m of the gene copies enter from other populations, and the frequency is p
 Homogenizes the populations within a species (unopposed by other forces) Rate of gene flow = m Rate of gene flow = m A 1 varies among populations (p i ) resident A 1 varies among populations (p i ) resident p i and the average allele frequency is p (source) p i and the average allele frequency is p (source) within population i (p i ), a proportion of m of the gene copies enter from other populations, and the frequency is p within population i (p i ), a proportion of m of the gene copies enter from other populations, and the frequency is p Homogenizes the populations within a species (unopposed by other forces) Homogenizes the populations within a species (unopposed by other forces) Rate of gene flow = m Rate of gene flow = m A 1 varies among populations (p i ) resident A 1 varies among populations (p i ) resident p i and the average allele frequency is p (source) p i and the average allele frequency is p (source) within population i (p i ), a proportion of m of the gene copies enter from other populations, and the frequency is p within population i (p i ), a proportion of m of the gene copies enter from other populations, and the frequency is p")
44
Gene flow pp m Source resident

45
Gene flow A proportion 1-m of the gene copies are non-immigrants and among these the frequency is p A proportion 1-m of the gene copies are non-immigrants and among these the frequency is p After 1 generation the populations new allele freq (p’) is: After 1 generation the populations new allele freq (p’) is: p’ = p(1-m) + pm or p’ = p-pm + pm So........Δp = m(p – p) A proportion 1-m of the gene copies are non-immigrants and among these the frequency is p A proportion 1-m of the gene copies are non-immigrants and among these the frequency is p After 1 generation the populations new allele freq (p’) is: After 1 generation the populations new allele freq (p’) is: p’ = p(1-m) + pm or p’ = p-pm + pm So........Δp = m(p – p)
 is: After 1 generation the populations new allele freq (p’) is: p’ = p(1-m) + pm or p’ = p-pm + pm So Δp = m(p – p) A proportion 1-m of the gene copies are non-immigrants and among these the frequency is p A proportion 1-m of the gene copies are non-immigrants and among these the frequency is p After 1 generation the populations new allele freq (p’) is: After 1 generation the populations new allele freq (p’) is: p’ = p(1-m) + pm or p’ = p-pm + pm So Δp = m(p – p)")
46
Gene flow p=0.3p=0.1 m=0.001 Source resident 1 in a 1000 is a migrant

47
Gene flow Δp = m(p-p) = 0.001(0.3-0.1) =.0002 p’ = p(1-m) + pm = 0.1(1-0.001)+0.3*0.001 =0.1002 Δp = m(p-p) = 0.001(0.3-0.1) =.0002 p’ = p(1-m) + pm = 0.1(1-0.001)+0.3*0.001 =0.1002
 = 0.001( ) =.0002 p’ = p(1-m) + pm = 0.1( )+0.3*0.001 = Δp = m(p-p) = 0.001( ) =.0002 p’ = p(1-m) + pm = 0.1( )+0.3*0.001 =0.1002")
48
Equilibrium Frequency Equilibrium frequency is found by setting p’ = 0 (or Δp = 0), which is where p’ is not changing between generations and is, thus, in equilibrium Equilibrium frequency is found by setting p’ = 0 (or Δp = 0), which is where p’ is not changing between generations and is, thus, in equilibrium So... Δp = 0 = m(p-p) thus p = p Therefore, each population will ultimately attain the same allele frequencies showing that gene flow homogenizes populations Equilibrium frequency is found by setting p’ = 0 (or Δp = 0), which is where p’ is not changing between generations and is, thus, in equilibrium Equilibrium frequency is found by setting p’ = 0 (or Δp = 0), which is where p’ is not changing between generations and is, thus, in equilibrium So... Δp = 0 = m(p-p) thus p = p Therefore, each population will ultimately attain the same allele frequencies showing that gene flow homogenizes populations
 thus p = p Therefore, each population will ultimately attain the same allele frequencies showing that gene flow homogenizes populations Equilibrium frequency is found by setting p’ = 0 (or Δp = 0), which is where p’ is not changing between generations and is, thus, in equilibrium Equilibrium frequency is found by setting p’ = 0 (or Δp = 0), which is where p’ is not changing between generations and is, thus, in equilibrium So... Δp = 0 = m(p-p) thus p = p Therefore, each population will ultimately attain the same allele frequencies showing that gene flow homogenizes populations.")
49
Gene flow and drift In the absence of gene flow populations tend to diverge due to drift In the absence of gene flow populations tend to diverge due to drift F st = fraction of autozygotes in a sub-population at time t F st = fraction of autozygotes in a sub-population at time t Another way to look at this is that F st is a measure of the observed variation in allele frequency among populations Another way to look at this is that F st is a measure of the observed variation in allele frequency among populations F st = 1- (1-½N) t see page 315, box 11.D In the absence of gene flow populations tend to diverge due to drift In the absence of gene flow populations tend to diverge due to drift F st = fraction of autozygotes in a sub-population at time t F st = fraction of autozygotes in a sub-population at time t Another way to look at this is that F st is a measure of the observed variation in allele frequency among populations Another way to look at this is that F st is a measure of the observed variation in allele frequency among populations F st = 1- (1-½N) t see page 315, box 11.D
 t see page 315, box 11.D In the absence of gene flow populations tend to diverge due to drift In the absence of gene flow populations tend to diverge due to drift F st = fraction of autozygotes in a sub-population at time t F st = fraction of autozygotes in a sub-population at time t Another way to look at this is that F st is a measure of the observed variation in allele frequency among populations Another way to look at this is that F st is a measure of the observed variation in allele frequency among populations F st = 1- (1-½N) t see page 315, box 11.D")
50
Gene flow and Drift F st must reach an equilibrium between drift and gene flow F st must reach an equilibrium between drift and gene flow Thus F st = 1 / (4Nm + 1) Thus F st = 1 / (4Nm + 1) F st must reach an equilibrium between drift and gene flow F st must reach an equilibrium between drift and gene flow Thus F st = 1 / (4Nm + 1) Thus F st = 1 / (4Nm + 1)
 Thus F st = 1 / (4Nm + 1) F st must reach an equilibrium between drift and gene flow F st must reach an equilibrium between drift and gene flow Thus F st = 1 / (4Nm + 1) Thus F st = 1 / (4Nm + 1)")
51
Gene flow We can rearrange F st = 1 / (4Nm + 1) so that we can estimate the average # of immigrants into a population per generation We can rearrange F st = 1 / (4Nm + 1) so that we can estimate the average # of immigrants into a population per generation Nm = 1 / (F st – 1) Notice! This tells us that the higher the rate of gene flow, the more similar the allele frequencies between populations Notice! This tells us that the higher the rate of gene flow, the more similar the allele frequencies between populations Conversely, observing a strong divergence indicates that the balance between gene flow and drift is tipped toward drift Conversely, observing a strong divergence indicates that the balance between gene flow and drift is tipped toward drift We can rearrange F st = 1 / (4Nm + 1) so that we can estimate the average # of immigrants into a population per generation We can rearrange F st = 1 / (4Nm + 1) so that we can estimate the average # of immigrants into a population per generation Nm = 1 / (F st – 1) Notice! This tells us that the higher the rate of gene flow, the more similar the allele frequencies between populations Notice! This tells us that the higher the rate of gene flow, the more similar the allele frequencies between populations Conversely, observing a strong divergence indicates that the balance between gene flow and drift is tipped toward drift Conversely, observing a strong divergence indicates that the balance between gene flow and drift is tipped toward drift
 so that we can estimate the average # of immigrants into a population per generation We can rearrange F st = 1 / (4Nm + 1) so that we can estimate the average # of immigrants into a population per generation Nm = 1 / (F st – 1) Notice. This tells us that the higher the rate of gene flow, the more similar the allele frequencies between populations Notice. This tells us that the higher the rate of gene flow, the more similar the allele frequencies between populations Conversely, observing a strong divergence indicates that the balance between gene flow and drift is tipped toward drift Conversely, observing a strong divergence indicates that the balance between gene flow and drift is tipped toward drift.")
52
Estimating F st Direct estimates of gene flow Direct estimates of gene flow Mark-recapture studies... Mark-recapture studies... Indirect estimates of gene flow Indirect estimates of gene flow Alleles used to calculate F st are neutral Alleles used to calculate F st are neutral Allele frequencies must have reached equilibrium between gene flow and drift Allele frequencies must have reached equilibrium between gene flow and drift Direct estimates of gene flow Direct estimates of gene flow Mark-recapture studies... Mark-recapture studies... Indirect estimates of gene flow Indirect estimates of gene flow Alleles used to calculate F st are neutral Alleles used to calculate F st are neutral Allele frequencies must have reached equilibrium between gene flow and drift Allele frequencies must have reached equilibrium between gene flow and drift

53
Gene Flow Example! Water snakes of Lake Erie Nerodia sipedon Live on mainland and on several islands Color pattern variable Strongly banded to unbanded Banding controlled by a single locus with two alleles Banded is dominant over unbanded On mainland most snakes are banded On island many are unbanded Water snakes of Lake Erie Nerodia sipedon Live on mainland and on several islands Color pattern variable Strongly banded to unbanded Banding controlled by a single locus with two alleles Banded is dominant over unbanded On mainland most snakes are banded On island many are unbanded

55
Gene Flow... Water snakes of Lake Erie Banding pattern due to natural selection On islands snakes bask on rocks On mainland stay closer to vegetation Why is the unbanded allele not fixed on islands? Banded snakes migrate from mainland each generation Bring banded alleles to island gene pool Migration works in opposition to natural selection Water snakes of Lake Erie Banding pattern due to natural selection On islands snakes bask on rocks On mainland stay closer to vegetation Why is the unbanded allele not fixed on islands? Banded snakes migrate from mainland each generation Bring banded alleles to island gene pool Migration works in opposition to natural selection

57
Conservation Genetics Westermeier’s hypothesis Westermeier’s hypothesis Destruction of prairie did two things Destruction of prairie did two things Reduced population size Fragmented remaining population Remaining prairie chickens were trapped in two islands in seas of farmland Remaining prairie chickens were trapped in two islands in seas of farmland Genetic drift caused decline in heterozygosity Genetic drift caused decline in heterozygosity Inbreeding depression occurred Inbreeding depression occurred Westermeier’s hypothesis Westermeier’s hypothesis Destruction of prairie did two things Destruction of prairie did two things Reduced population size Fragmented remaining population Remaining prairie chickens were trapped in two islands in seas of farmland Remaining prairie chickens were trapped in two islands in seas of farmland Genetic drift caused decline in heterozygosity Genetic drift caused decline in heterozygosity Inbreeding depression occurred Inbreeding depression occurred

58
Conservation Genetics Accumulation of deleterious recessives leads to reduction in population size Accumulation of deleterious recessives leads to reduction in population size Effectiveness of genetic drift is increased Effectiveness of genetic drift is increased Speed and proportion of deleterious mutations going to fixation increases Speed and proportion of deleterious mutations going to fixation increases Population size decreases more Population size decreases more Mutational Meltdown Mutational Meltdown Accumulation of deleterious recessives leads to reduction in population size Accumulation of deleterious recessives leads to reduction in population size Effectiveness of genetic drift is increased Effectiveness of genetic drift is increased Speed and proportion of deleterious mutations going to fixation increases Speed and proportion of deleterious mutations going to fixation increases Population size decreases more Population size decreases more Mutational Meltdown Mutational Meltdown

60
Conservation Genetics Prairie chickens caught in mutational meltdown Prairie chickens caught in mutational meltdown Reproductive success decreased Reproductive success decreased Hatching success low Hatching success low Birds had fallen into extinction vortex Birds had fallen into extinction vortex Birds needed gene flow Birds needed gene flow Prairie chickens caught in mutational meltdown Prairie chickens caught in mutational meltdown Reproductive success decreased Reproductive success decreased Hatching success low Hatching success low Birds had fallen into extinction vortex Birds had fallen into extinction vortex Birds needed gene flow Birds needed gene flow

61
Conservation Genetics In 1992 conservationists trapped birds from Minnesota, Kansas, and Nebraska and moved them to Jasper County, Illinois In 1992 conservationists trapped birds from Minnesota, Kansas, and Nebraska and moved them to Jasper County, Illinois Hatching rate increased and population began to grow Hatching rate increased and population began to grow Migration, genetic drift, and nonrandom mating all contributed to fate of Illinois greater prairie chickens Migration, genetic drift, and nonrandom mating all contributed to fate of Illinois greater prairie chickens In 1992 conservationists trapped birds from Minnesota, Kansas, and Nebraska and moved them to Jasper County, Illinois In 1992 conservationists trapped birds from Minnesota, Kansas, and Nebraska and moved them to Jasper County, Illinois Hatching rate increased and population began to grow Hatching rate increased and population began to grow Migration, genetic drift, and nonrandom mating all contributed to fate of Illinois greater prairie chickens Migration, genetic drift, and nonrandom mating all contributed to fate of Illinois greater prairie chickens

62
Conclusions about Gene Flow 1.Certain taxa display substantial gene flow over long distances 2.The average level of gene flow is greatly restricted over short distances Implies local populations can diverge substantially due to drift Implies local populations can diverge substantially due to drift Thus, species can adapt to local conditions!!!! Thus, species can adapt to local conditions!!!! 1.Certain taxa display substantial gene flow over long distances 2.The average level of gene flow is greatly restricted over short distances Implies local populations can diverge substantially due to drift Implies local populations can diverge substantially due to drift Thus, species can adapt to local conditions!!!! Thus, species can adapt to local conditions!!!!

Similar presentations













>")

 that mendel described.>")




>")




