Download presentation
Presentation is loading. Please wait.

1
Molecular Motors Filament…….motor Actin…….….myosins Tubulin….….dynein and kinesins

2
Myosin Family Tree Is Large and Diverse

3
Differences in the myosin tails influence movement along the actin cytoskeleton

4
Kinesin and Dynein Comprise 2 classes of MT motors Kinesins are involved in organelle transport, in mitosis, in meiosis, and in the transport of synaptic vesicles along axons. Cytoplasmic dyneins are involved in organelle transport and mitosis. Organelles and vesicles containing kinesin move from the minus end of a microtubule (at a microtubule organizing center, such as centrosome) to the plus end (from the center of a cell to its periphery, called anterograde transport). Cytoplasmic dynein moves the particles from the plus end to the minus end of the microtubules (retrograde transport). Ciliary dynein is a much larger protein than the cytoplasmic dynein: its mass is about 2 million daltons, composed of either 2 or 3 heavy chains and 10 or more smaller polypeptides. The dynein motor protein in cilia and flagella has a head which hydrolyzes ATP and interacts with the adjacent microtubules to generate a sliding force between the microtubules. However, the microtubules are linked together, therefore, they can not slide but must bend. This local bending of the microtubules is the mechanism of ciliary movement, the beating.
 to the plus end (from the center of a cell to its periphery, called anterograde transport). Cytoplasmic dynein moves the particles from the plus end to the minus end of the microtubules (retrograde transport). Ciliary dynein is a much larger protein than the cytoplasmic dynein: its mass is about 2 million daltons, composed of either 2 or 3 heavy chains and 10 or more smaller polypeptides. The dynein motor protein in cilia and flagella has a head which hydrolyzes ATP and interacts with the adjacent microtubules to generate a sliding force between the microtubules. However, the microtubules are linked together, therefore, they can not slide but must bend. This local bending of the microtubules is the mechanism of ciliary movement, the beating..")
5
The structural similarity between myosin and kinesin head groups indicates a common evolutionary origin

7
Microtubules are polymers of and tubulin GTP-binding heterodimers that form a ring of 13 monomers

8
Microtubules grow faster at the plus end than the minus end

10
Cell was injected with rhodamine-tubulin such that 1 out of 20 tubulins was fluorescent. Using a “dark mark” in the center of the microtubule as A reference, it’s apparent that the plus end grows and The minus end shrinks. (Rather than the microtubule Moving from left to right). The plus end also shows dynamic Instability.
. The plus end also shows dynamic Instability..")
11
Dynamic Instability allows Transient MT Growth

14
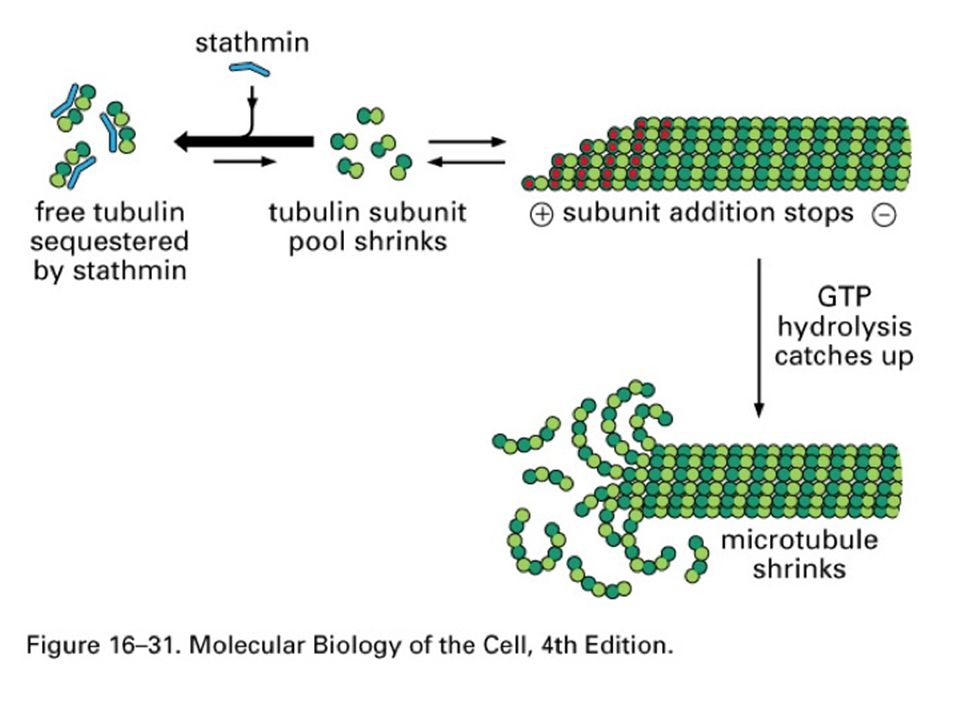
Catastrophe is loss of GTP end (Occurs when GTP hydrolysis exceeds binding) Rescue is binding of tubulin GTP onto end
 Rescue is binding of tubulin GTP onto end")
15
Microtubule Associated Proteins (MAPs)
")
16
-tubulin ring complex ( -TuRC) nucleates mictrtubule assembly It’s at the MINUS end.
 nucleates mictrtubule assembly It’s at the MINUS end.")
17
The effects of proteins that bind to microtubule ends MAP kinase = mitogen activated protein MAP = microtubule associated proteins

18
Katanin (Japanese for sword) is a microtubule Severing protein Taxol-stabilized rhodamine-labelled microtubules were fixed to glass slides. Purified katanin and ATP were added; some breaks seen after 30 sec; many breaks 3 min.

19
Drugs can influence cytoskeleton integrity (can stabilize or destabilize)
")
20
Chemical Synthesis of Taxol is a recent advance; used to treat cancer

21
Microtubule Functions

22
Centrioles (Basal Bodies) Promote Mitosis Microtubules can form Cilia (Flagella) and basal bodies (centrioles)
 Promote Mitosis Microtubules can form Cilia (Flagella) and basal bodies (centrioles)")
23
The center of the center of the cell… Centrioles in a centrosome connect to centromeres MTOC=microtubule organizing center

24
Centromeres bound to microtubules bound to centrosomes

25
-Small square wells were micromachined into a plastic substrate -A single centrosome was placed inside along with depolymerized Microtubules -As microtubules polymerize they push against the walls of the square -This leads to a centering effect on the centrosome Self-organization as a Guiding Principle in Polarity

26
A GTPase can act as a MT directionality signal The RanGTPase cycle provides directionality to nucleocytoplasmic transport, regulating interactions between cargoes and nuclear transport receptors of the importin-beta family. The Ran-importin-beta system also functions in mitotic spindle assembly and nuclear pore and nuclear envelope formation. The common principle underlying these diverse functions throughout the cell cycle is thought to be anisotropy of the distribution of RanGTP (the RanGTP gradient), driven by the chromatin-associated guanine nucleotide exchange factor RCC1. However, the existence and function of a RanGTP gradient during mitosis in cells is unclear. Here we examine the Ran–importin-beta system in cells by conventional and fluorescence lifetime microscopy using a biosensor, termed Rango, that increases its fluorescence resonance energy transfer signal when released from importin-beta by RanGTP. Rango is predominantly free in mitotic cells, but is further liberated around mitotic chromatin. In vitro experiments and modelling show that this localized increase of free cargoes corresponds to changes in RanGTP concentration sufficient to stabilize microtubules in extracts. In cells, the Ran–importin-beta–cargo gradient kinetically promotes spindle formation but is largely dispensable once the spindle has been established. Consistent with previous reports, we observe that the Ran system also affects spindle pole formation and chromosome congression in vivo.
, driven by the chromatin-associated guanine nucleotide exchange factor RCC1. However, the existence and function of a RanGTP gradient during mitosis in cells is unclear. Here we examine the Ran–importin-beta system in cells by conventional and fluorescence lifetime microscopy using a biosensor, termed Rango, that increases its fluorescence resonance energy transfer signal when released from importin-beta by RanGTP. Rango is predominantly free in mitotic cells, but is further liberated around mitotic chromatin. In vitro experiments and modelling show that this localized increase of free cargoes corresponds to changes in RanGTP concentration sufficient to stabilize microtubules in extracts. In cells, the Ran–importin-beta–cargo gradient kinetically promotes spindle formation but is largely dispensable once the spindle has been established. Consistent with previous reports, we observe that the Ran system also affects spindle pole formation and chromosome congression in vivo..")
27
A GTPase can act as a MT directionality signal

Similar presentations














 * Cancer diseases therapy: taxanes.>")

>")




 E. D. Salmon Biology Reading: Lodish et al., Molecular Cell Biology; Alberts et al., Molecular Biology of the.>")
