Download presentation
Presentation is loading. Please wait.

1
SEMINARS!!!! Dr. Andy Baldwin, Department of Biochemistry, University of Toronto. Title: "The structure and dynamics of AlphaB-crystallin oligomers, determined using solution-state nuclear magnetic resonance spectroscopy, ion-mobility mass spectrometry and electron microscopy" F 3:15 pm.

2
Exam R 6-8 pm Sl 110

4
Glygogen Muscle: 1-2% Liver: 10%!! Breakdown Synthesis

8
Figure 15-28 G6 Phosphatase is located on ER membrane.

9
Synthesis

12
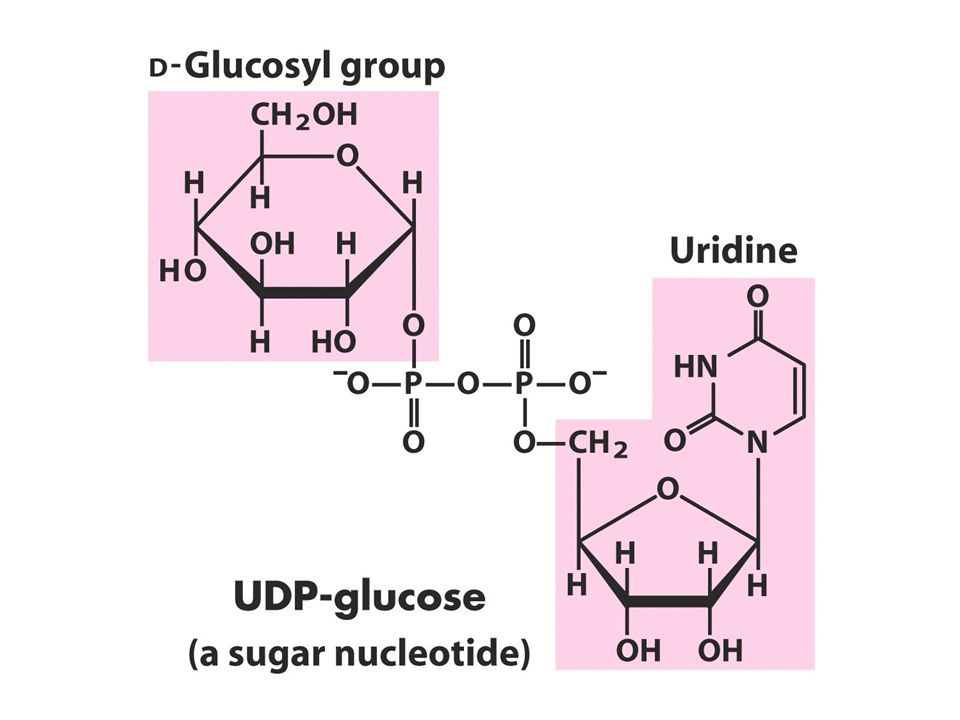
Nucleotide donors: formation irreversible good for enzyme recognition are excellent leaving groups

13
Glycogenin catalyzes two distinct reactions. Initial attack by the hydroxyl group of Tyr194 on C-1 of the glucosyl moiety of UDP-glucose results in a glucosylated Tyr residue. The C-1 of another UDP-glucose molecule is now attacked by the C-4 hydroxyl group of the terminal glucose, and this sequence repeats to form a nascent glycogen molecule of eight glucose residues attached by (α1→4) glycosidic linkages.
 glycosidic linkages..")
14
Glycogenin structure. Muscle glycogenin (Mr 37,000) forms dimers in solution. The substrate, UDP-glucose, is bound to a Rossmann fold near the amino terminus and is some distance from the Tyr194 residues—15 Å from the Tyr in the same monomer, 12 Å from the Tyr in the dimeric partner. Each UDP-glucose is bound through its phosphates to a Mn2+ ion that is essential to catalysis. Mn2+ is believed to function as an electron-pair acceptor to stabilize the leaving group, UDP. The glycosidic bond in the product has the same configuration about the C-1 of glucose as the substrate UDP-glucose, suggesting that the transfer of glucose from UDP to Tyr194 occurs in two steps. The first step is probably a nucleophilic attack by Asp162, forming a temporary intermediate with inverted configuration. A second nucleophilic attack by Tyr194 then restores the starting configuration

16
Glycogenin and the structure of the glycogen particle. (b) Structure of the glycogen particle. Starting at a central glycogenin molecule, glycogen chains (12 to 14 residues) extend in tiers. Inner chains have two (α1→6) branches each. Chains in the outer tier are unbranched. Particles consist of about 55,000 glucose residues in a molecule of about 21 nm diameter and Mr ~1 × 107.
 Structure of the glycogen particle. Starting at a central glycogenin molecule, glycogen chains (12 to 14 residues) extend in tiers. Inner chains have two (α1→6) branches each. Chains in the outer tier are unbranched. Particles consist of about 55,000 glucose residues in a molecule of about 21 nm diameter and Mr ~1 ×")
17
Regulation of muscle glycogen phosphorylase by covalent modification.

18
Cascade mechanism of epinephrine and glucagon action.

20
Figure 18-22The enzymatic activities of phosphorylase a and glycogen synthase in mouse liver in response to an infusion of glucose. Page 648

21
Figure 18-9The control of glycogen phosphorylase activity. Page 635

22
Page 639 Figure 18-13 Control of glycogen metabolism in muscle.

23
Figure 18-16X-Ray structure of rat testis calmodulin. Page 642

24
Figure 18-19 Schematic diagram of the Ca 2+ –CaM-dependent activation of protein kinases.

25
Figure 18-21The antagonistic effects of insulin and epinephrine on glycogen metabolism in muscle. Page 645

26
Maintaining Blood Glucose Levels During exercise or long after meals, the liver releases glc into the bloodstream Glc inhibits pancreatic -cells from secreting glucagon. Inhibition is released when glc levels fall. Glucagon receptors on liver cells respond to glucagon binding by activating AC causing [cAMP]. [cAMP] increases the rate of glycogen breakdown and increased G6P. G6P cannot pass through cell membranes. However, the liver, which doesn’t rely on glc for a major energy source, has a G6P hydrolase to release glc.

27
Figure 18-24Formation and degradation of -D- fructose-2,6-bisphosphate as catalyzed by PFK-2 and FBPase-2. Page 649

28
Figure 18-26a The liver’s response to stress. (a) Stimulation of -adrenoreceptors by epinephrine activates phospholipase C to hydrolyze PIP 2 to IP 3 and DAG. Page 652
 Stimulation of -adrenoreceptors by epinephrine activates phospholipase C to hydrolyze PIP 2 to IP 3 and DAG. Page 652.")
29
Figure 18-26b The liver’s response to stress. (b) The participation of two second messenger systems. Page 652
 The participation of two second messenger systems. Page 652.")
30
Effects of glycogen synthase kinase 3 (GSK3) on glycogen synthase activity.
 on glycogen synthase activity.")
31
phosphoprotein phosphatase 1 (PP1) Phosphatidylinositol 3-kinase (PI-3K) converts phosphatidylinositol 4,5-bisphosphate (PIP2) in the membrane to phosphatidylinositol 3,4,5-trisphosphate (PIP3). insulin receptor substrate-1 (IRS-1) glycogen synthase kinase 3 (GSK3)
 glycogen synthase kinase 3 (GSK3).")
32
The glycogen-targeting protein GM is one of a family of proteins that bind other proteins (including PP1) to glycogen particles.
 to glycogen particles.")
35
Glycogen phosphorylase of liver as a glucose sensor.

36
FIGURE 15-36 (part 1) Glycogen phosphorylase of liver as a glucose sensor. Glucose binding to an allosteric site of the phosphorylase a isozyme of liver induces a conformational change that exposes its phosphorylated Ser residues to the action of phosphorylase a phosphatase (PP1). This phosphatase converts phosphorylase a to phosphorylase b, sharply reducing the activity of phosphorylase and slowing glycogen breakdown in response to high blood glucose. Insulin also acts indirectly to stimulate PP1 and slow glycogen breakdown.
. This phosphatase converts phosphorylase a to phosphorylase b, sharply reducing the activity of phosphorylase and slowing glycogen breakdown in response to high blood glucose. Insulin also acts indirectly to stimulate PP1 and slow glycogen breakdown..")
37
FIGURE 15-36 (part 2) Glycogen phosphorylase of liver as a glucose sensor. Glucose binding to an allosteric site of the phosphorylase a isozyme of liver induces a conformational change that exposes its phosphorylated Ser residues to the action of phosphorylase a phosphatase (PP1). This phosphatase converts phosphorylase a to phosphorylase b, sharply reducing the activity of phosphorylase and slowing glycogen breakdown in response to high blood glucose. Insulin also acts indirectly to stimulate PP1 and slow glycogen breakdown.
. This phosphatase converts phosphorylase a to phosphorylase b, sharply reducing the activity of phosphorylase and slowing glycogen breakdown in response to high blood glucose. Insulin also acts indirectly to stimulate PP1 and slow glycogen breakdown..")
38
LEHNINGER PRINCIPLES OF BIOCHEMISTRY Fifth Edition David L. Nelson and Michael M. Cox © 2008 W. H. Freeman and Company CHAPTER 16 The Citric Acid Cycle

41
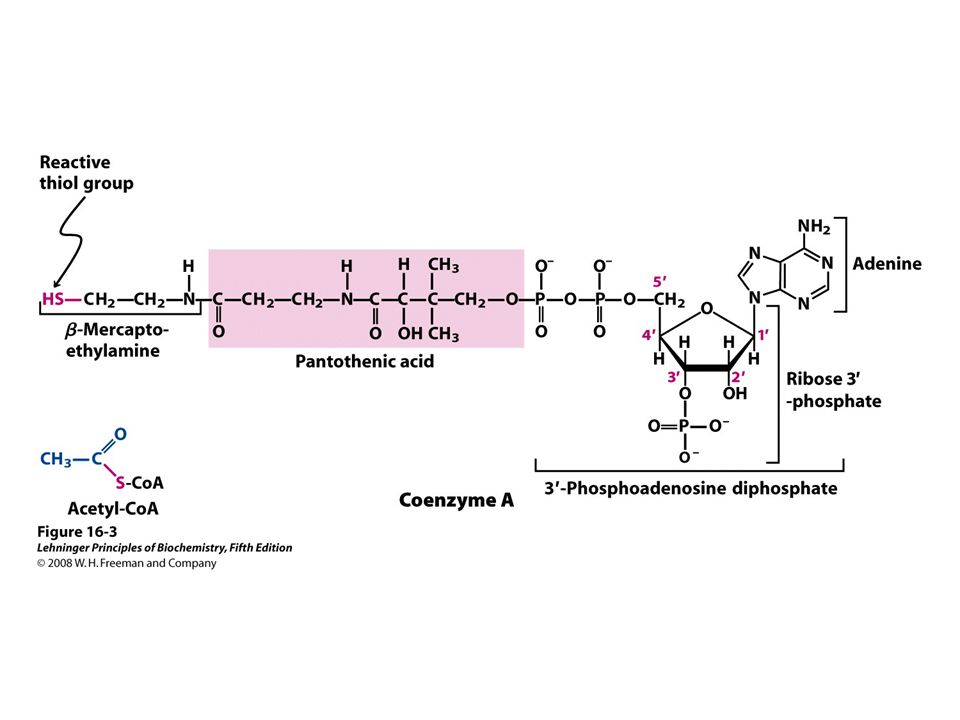
AcetylCo A + 3 NAD + + FAD + GDP + P i 2 CO 2 + 3 NADH + FADH 2 + GTP + CoA TCA Cycle : Pyruvate + Coenzyme A + NAD + Acetyl CoA + CO 2 + NADH Pyruvate Dehyrdogenase Reaction:

42
Figure 17-3a PDH complex

43
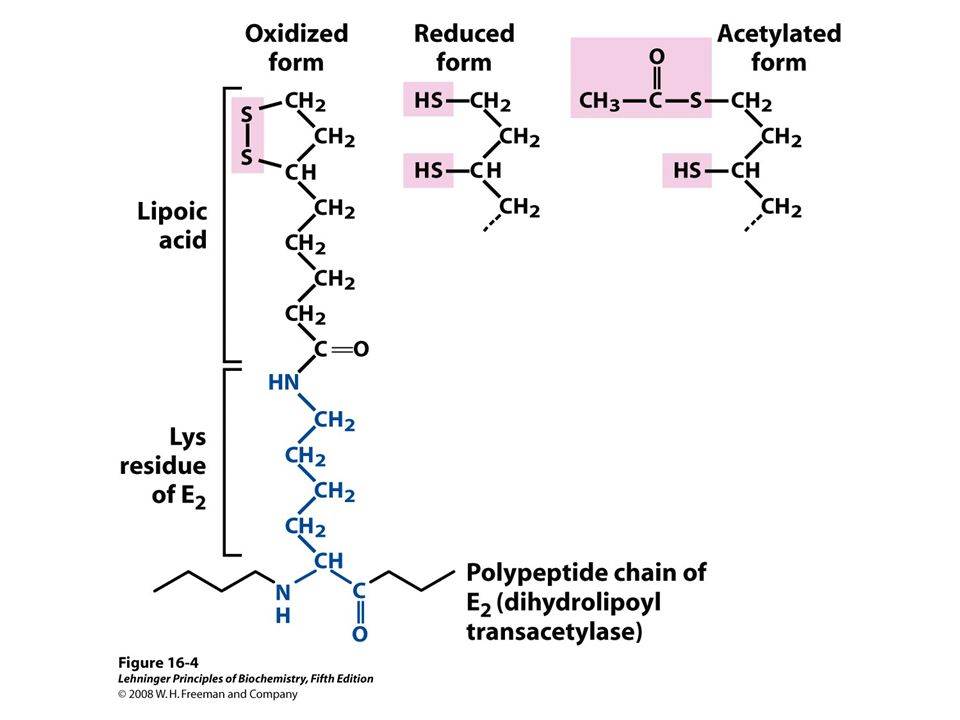
Figure 21-4 Structural organization of the E. coli PDC. Page 769 E2 Dihydrolypoly transacetlyase core (trimers) PDH: 24 Subunits (E1) (as dimers) 24 subunits 12 dihydrolypoyl dehydrogenase (E3) (as dimers) a+b Even more complex in yeast and mammals!
 PDH: 24 Subunits (E1) (as dimers) 24 subunits 12 dihydrolypoyl dehydrogenase (E3) (as dimers) a+b Even more complex in yeast and mammals!.")
44
Table 17-1

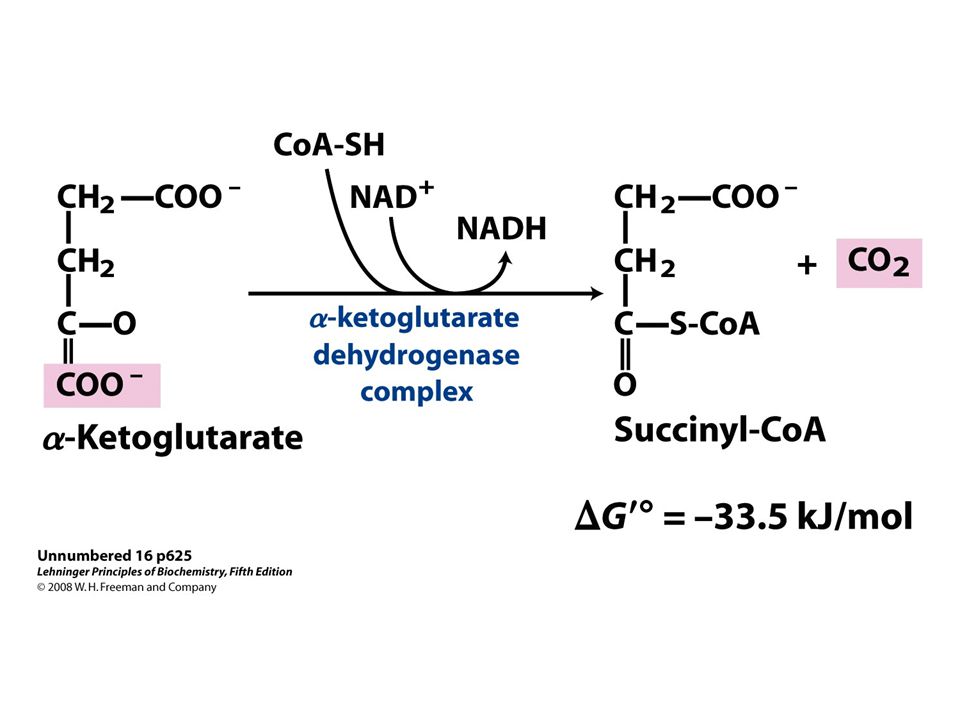
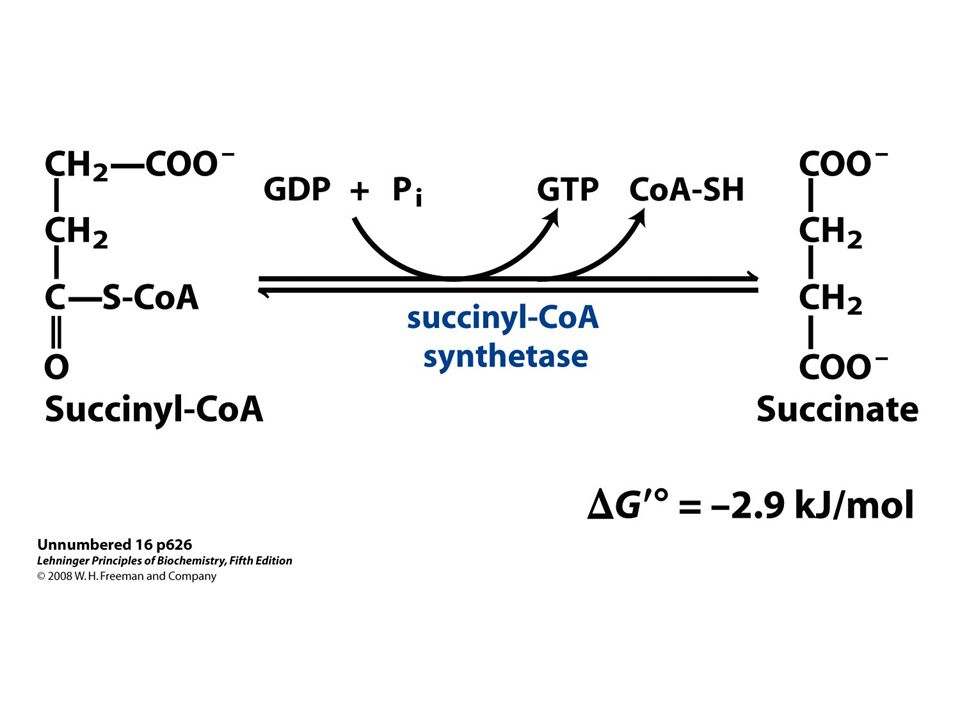
53
Structure of citrate synthase. The flexible domain of each subunit undergoes a large conformational change on binding oxaloacetate, creating a binding site for acetyl-CoA. (a) Open form of the enzyme alone (PDB ID 5CSC); (b) closed form with bound oxaloacetate and a stable analog of acetyl-CoA (carboxymethyl-CoA) (derived from PDB ID 5CTS).
 Open form of the enzyme alone (PDB ID 5CSC); (b) closed form with bound oxaloacetate and a stable analog of acetyl-CoA (carboxymethyl-CoA) (derived from PDB ID 5CTS)..")
58
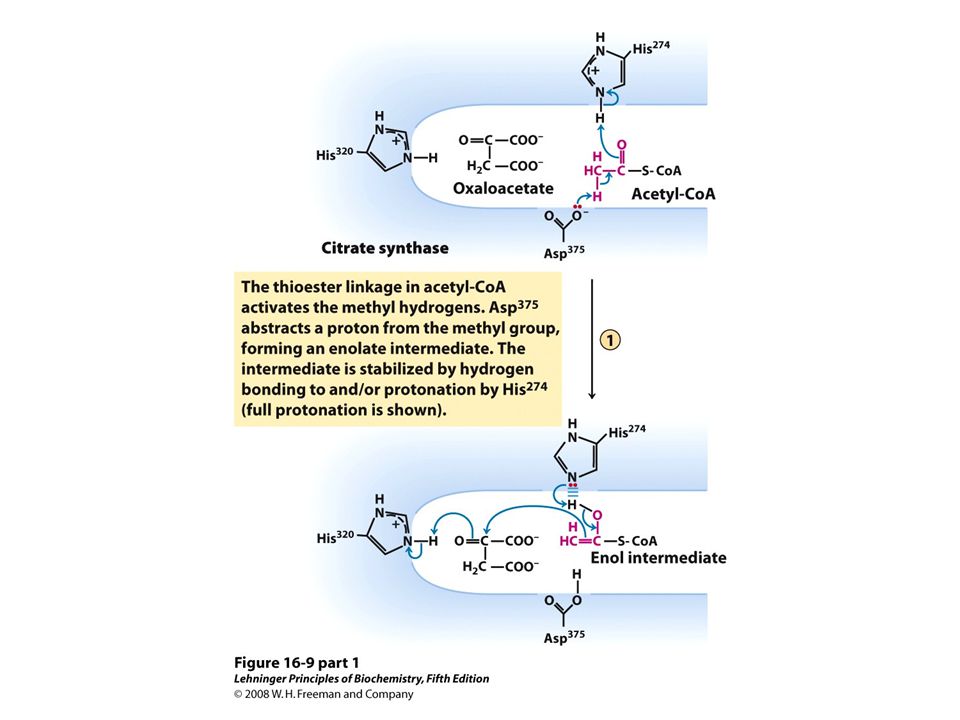
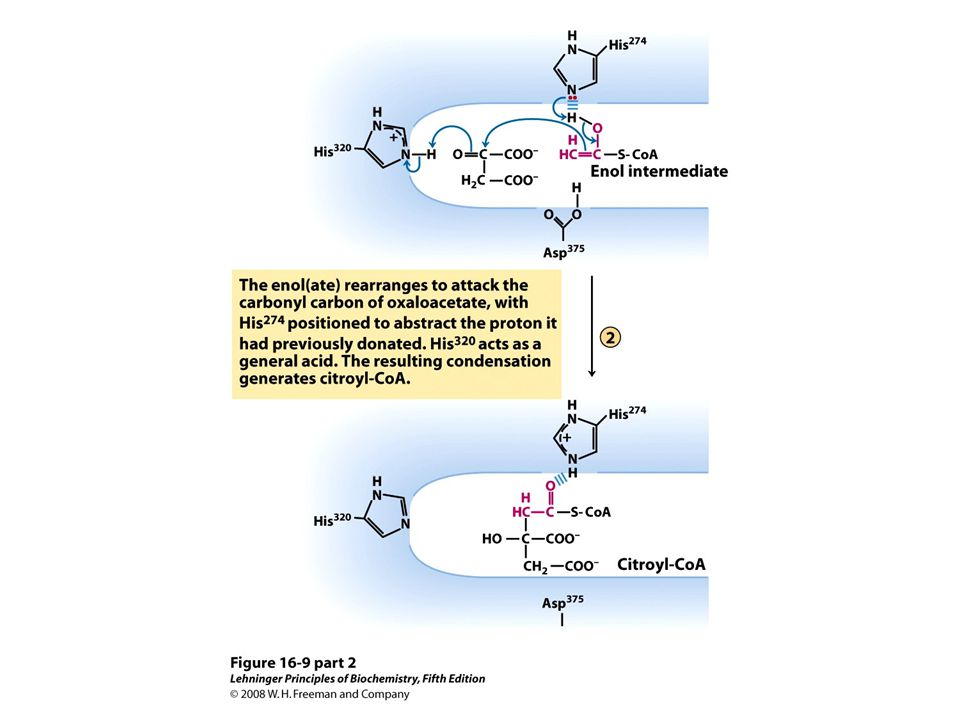
Citrate synthase mechanism http://bcs.whfreeman.com/lehninger5e/pages/bcs- main.asp?v=&s=16000&n=00010&i=16010.01&o=|00610|00580|00590|00510|00540|00600|0055 0|00570|00630|00010|00020|00030|00040|00070|00080|00090|00100|01000|02000|03000|04000| 05000|06000|07000|08000|09000|10000|11000|12000|13000|14000|15000|16000|17000|18000|1 9000|20000|21000|22000|23000|24000|25000|26000|27000|28000|99000|

82
“Alfonse, Biochemistry makes my head hurt!!” \

Similar presentations


























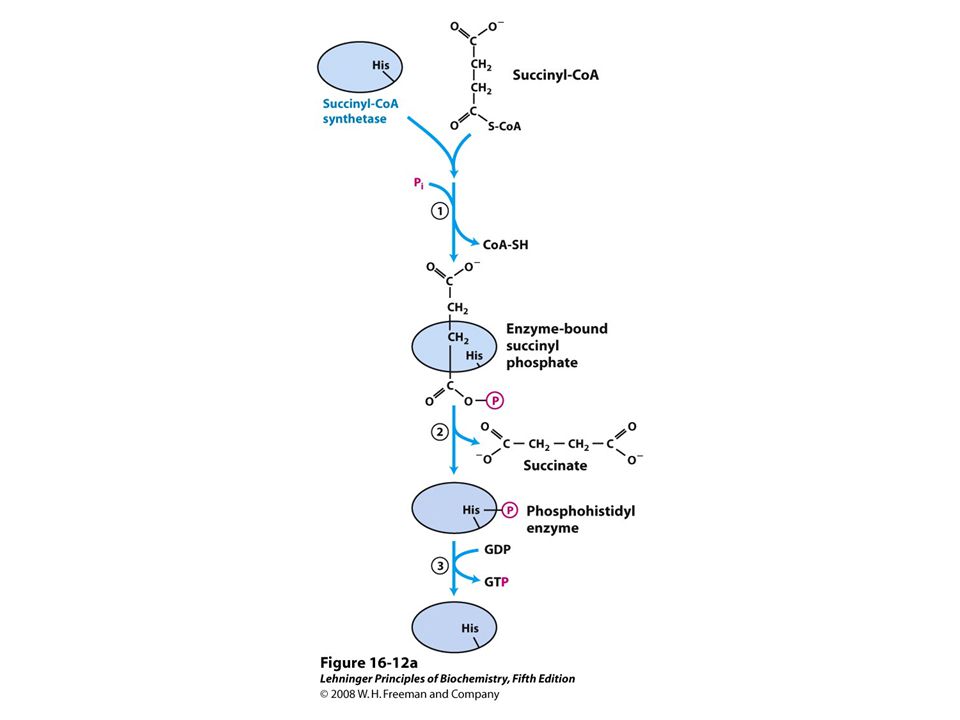


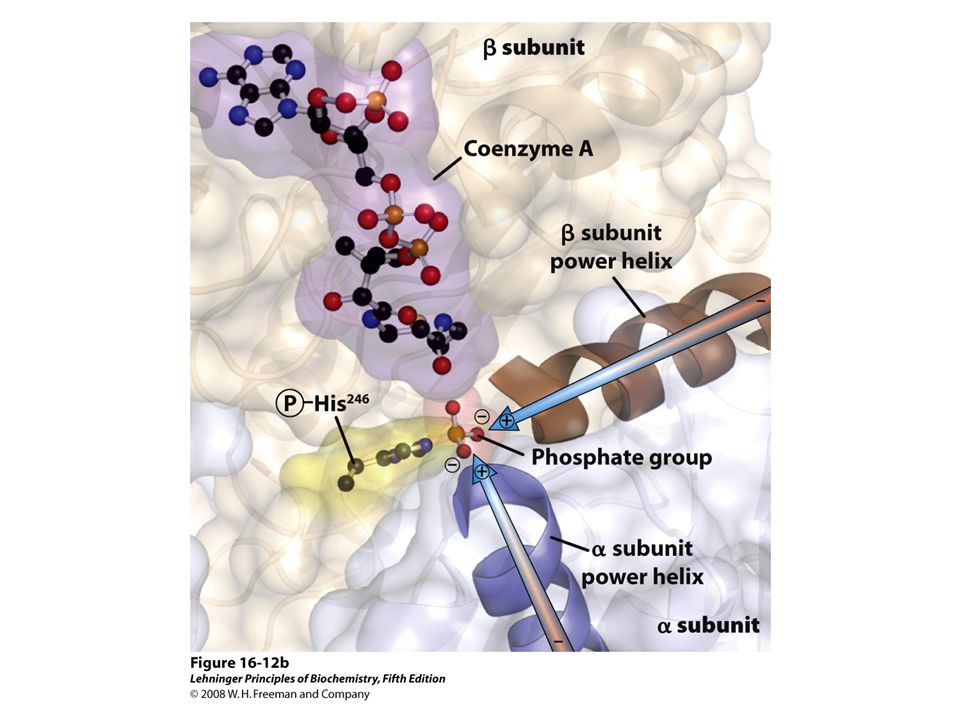


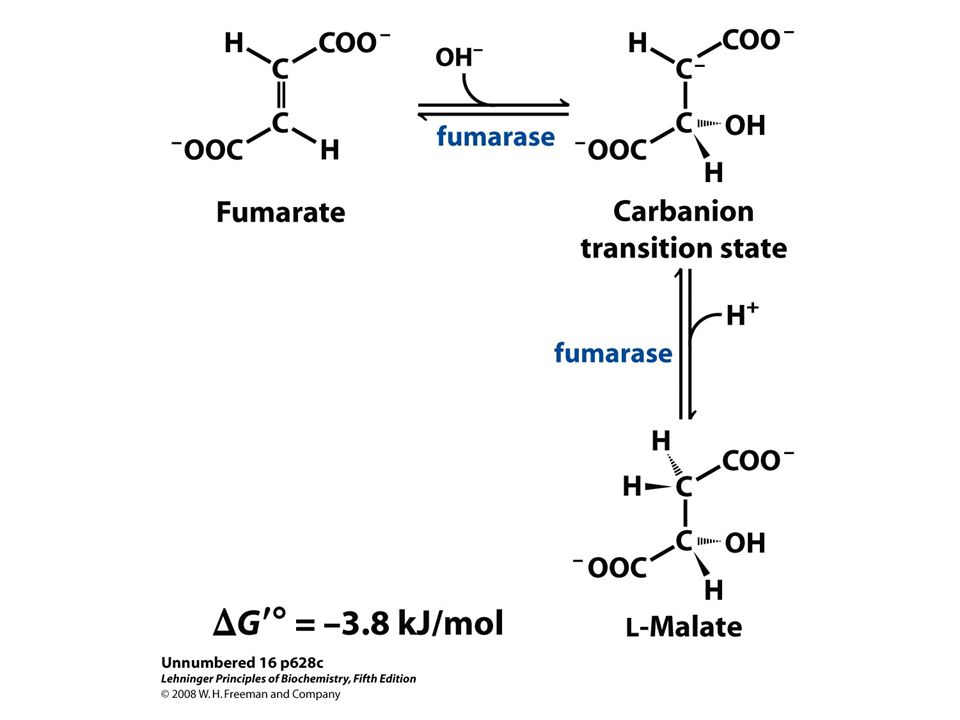
















 Molecular formula. (b) Schematic diagram illustrating its branched structure. Page 627.>")







 is amphibolic (both catabolic and anabolic) The cycle is involved in.>")
