Download presentation
1
Principles of Biochemistry
Horton • Moran • Scrimgeour • Perry • Rawn Principles of Biochemistry Fourth Edition Chapter 20 DNA Replication, Repair, and Recombination Copyright © 2006 Pearson Prentice Hall, Inc.

2
Chapter 20 - DNA Replication, Repair and Recombination
Holliday junction, an intermediate formed during recombination between two double-stranded DNA molecules

3
Fig 20.1 Semiconservative DNA replication
Each strand of DNA acts as a template for synthesis of a new strand Daughter DNA contains one parental and one newly synthesized strand

4
20.1 Chromosomal DNA Replication Is Bidirectional
E. coli chromosome is circular, double-stranded DNA (4.6x103 kilobase pairs, kb) Replication begins at a unique site (origin) Proceeds bidirectionally until the two replication complexes meet (termination site) Replisome - protein machinery for replication (one replisome at each of 2 replication forks)
 Replication begins at a unique site (origin) Proceeds bidirectionally until the two replication complexes meet (termination site) Replisome - protein machinery for replication (one replisome at each of 2 replication forks)")
5
Fig 20.2 Bidirectional DNA replication in E. coli
New strands of DNA are synthesized at the two replication forks where replisomes are located

6
Fig 20.2 (cont) Figure 20.2 Bidirectional DNA replication in Escherichia coli. Semiconservative DNA replication begins at a unique origin and proceeds in both directions. The synthesis of new strands of DNA (light gray) occurs at the two replication forks, where the replisomes are located. When the replication forks meet at the termination site, the two double-stranded DNA molecules separate. Note that each daughter molecule consists of one parental strand and one newly synthesized strand.
 occurs at the two replication forks, where the replisomes are located. When the replication forks meet at the termination site, the two double-stranded DNA molecules separate. Note that each daughter molecule consists of one parental strand and one newly synthesized strand.")
7
Fig. 20.3 Autoradiograph of a replicating E. coli chromosome
Figure 20.3 Autoradiograph of a replicating E. coli chromosome. The DNA was labeled with 3H-deoxythymidine, and the radioactivity detected by overlaying the replicating chromosome with photographic emulsion. The autoradiograph shows that the E. coli chromosome has two replication forks.

8
Eukaryotic Replication
Eukaryotic chromosomes are large linear, double-stranded DNA molecules Fruit fly large chromosomes ~5.0x104 kb (~10x larger than E. coli) Replication is bidirectional Multiple sites of initiation of DNA synthesis (versus one site in E. coli)
 Replication is bidirectional. Multiple sites of initiation of DNA synthesis (versus one site in E. coli)")
9
Fig 20.4 Replicating DNA in the fruit fly
Large number of replication forks at opposite ends of “bubbles” of duplicated DNA

10
20.2 DNA Polymerase E. coli contains three DNA polymerases
DNA polymerase I - repairs DNA and participates in DNA synthesis of one strand DNA polymerase II - role in DNA repair DNA polymerase III - the major DNA replication enzyme, responsible for chain elongation

12
Fig 20.5 DNA Polymerase III: subunit composition
Holoenzyme

13
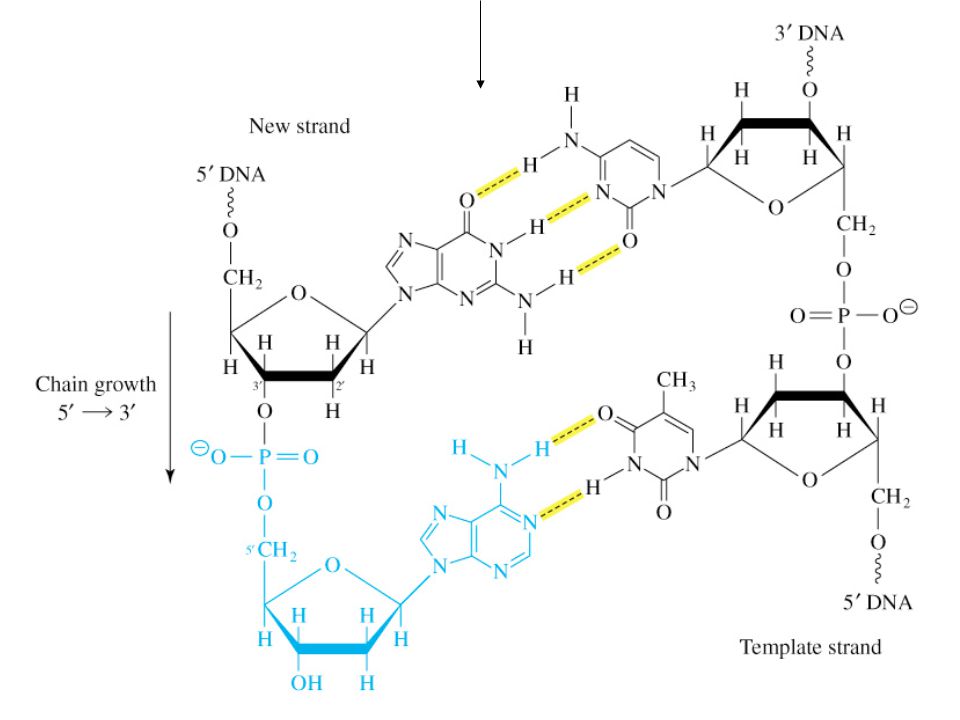
A. Chain Elongation Is a Nucleotidyl-Group-Transfer Reaction
Figure 20.6 (next 3 slides) Base pair between incoming deoxynucleotide 5’ triphosphate (blue) and a residue of the parental strand Terminal 3’ OH attacks a-phosphorous of incoming nucleotide to form new phosphodiester linkage
 Base pair between incoming deoxynucleotide 5’ triphosphate (blue) and a residue of the parental strand. Terminal 3’ OH attacks a-phosphorous of incoming nucleotide to form new phosphodiester linkage.")
14
Figure 20.6 Elongation of a DNA chain. During DNA synthesis, a base pair is created when an incoming deoxynucleoside 5'-triphosphate (blue) forms hydrogen bonds with a residue of the parental strand. A phosphodiester linkage forms when the terminal 3'-hydroxyl group attacks the -phosphorus atom of the incoming nucleotide. Hydrolysis of the released pyrophosphate makes the reaction irreversible.
 forms hydrogen bonds with a residue of the parental strand. A phosphodiester linkage forms when the terminal 3 -hydroxyl group attacks the -phosphorus atom of the incoming nucleotide. Hydrolysis of the released pyrophosphate makes the reaction irreversible.")
17
B. DNA Polymerase III Remains Bound to the Replication Fork
DNA polymerase III is a processive enzyme (remains bound to the replication fork until replication is complete) b-Subunits form a sliding clamp which surrounds the DNA molecule
 b-Subunits form a sliding clamp which surrounds the DNA molecule.")
18
Fig 20.7 Sliding clamp of b-subunits of DNA pol III
E coli archeabacteria bacteriophage T4 Two b-subunits associate to form a head-to-tail dimer in the shape of a ring that completely surrounds the DNA Remaining subunits of DNA pol III are bound to this structure

19
Fig 20.8 Bacteriophage DNA polymerase bound to DNA

20
C. Proofreading Corrects Polymerization Errors
DNA polymerase III holoenzyme also possesses 3’ 5’ exonuclease activity Pol III can catalyze both chain elongation and degradation Recognizes distortion in the DNA caused by incorrectly paired bases Exonuclease activity removes mispaired nucleotide before polymerization continues

21
20.3 DNA Polymerase Synthesizes Two Strands Simultaneously
DNA pol III catalyzes chain elongation only in the 5’ 3’ direction (antiparallel DNA strands) Leading strand - synthesized by polymerization in the same direction as fork movement Lagging strand - synthesized by polymerization in the opposite direction of fork movement Two core complexes of DNA pol III, one for leading, one for lagging strand
 Leading strand - synthesized by polymerization in the same direction as fork movement. Lagging strand - synthesized by polymerization in the opposite direction of fork movement. Two core complexes of DNA pol III, one for leading, one for lagging strand.")
22
Fig 19.13 Complementary base pairing and stacking in DNA

23
Fig 20.9 Diagram of the replication fork

24
A. Lagging-Strand Synthesis is Discontinuous
Leading strand is synthesized as one continuous polynucleotide (beginning at origin and ending at the termination site) Lagging strand is synthesized discontinuously in short pieces (Okazaki fragments) Pieces of the lagging strand are then joined by a separate reaction
 Lagging strand is synthesized discontinuously in short pieces (Okazaki fragments) Pieces of the lagging strand are then joined by a separate reaction.")
25
Fig 20.10 Demonstration of discontinuous DNA synthesis

26
Fig 20.10

27
Fig (cont) (From previous slide)
 (From previous slide)")
28
B. Each Okazaki Fragment Begins with an RNA Primer
Primosome is a complex containing primase enzyme which synthesizes short pieces of RNA at the replication fork (complementary to the lagging-strand template) DNA pol III uses the RNA primer to start the lagging-strand DNA synthesis Replisome - includes primosome, DNA pol III
 DNA pol III uses the RNA primer to start the lagging-strand DNA synthesis. Replisome - includes primosome, DNA pol III.")
29
Fig 20.11 Diagram of lagging-strand synthesis

30
C. Okazaki Fragments Are Joined by Action of DNA Polymerase I and DNA Ligase
Okazaki fragments are joined to produce a continuous strand of DNA in 3 steps: (1) Removal of the RNA primer (2) Synthesis of replacement DNA (3) Sealing of adjacent DNA fragments
 Removal of the RNA primer. (2) Synthesis of replacement DNA. (3) Sealing of adjacent DNA fragments.")
31
Fig 20.12 Klenow (large) fragment of DNA pol I, lacks 3’-5’ exonuclease activity Used for DNA synthesis

32
DNA Polymerase I Activities
The 5’ 3’ activity of DNA pol I removes the RNA primer at the beginning of each Okazaki fragment Synthesizes nick translation: polymerase activity synthesizes DNA in place of RNA

33
DNA Ligase Activity Catalyzes the formation of a phosphodiester linkage between 3’-hydroxyl and 5’-phosphate of adjacent Okazaki fragments Eukaryotic enzymes require ATP cosubstrate E. coli DNA ligase uses NAD+ as a cosubstrate

34
Fig 20.13 Joining of Okazaki fragments by DNA pol I and DNA ligase

35
Fig (continued)
")
36
Fig (continued)
")
37
Fig (continued)
")
38
Fig 20.14 Mechanism of DNA ligase in E. coli

39
Figure 20.14 Proposed mechanism of DNA ligase in E. coli. Using NAD+ as a cosubstrate, E. coli DNA ligase catalyzes the formation of a phosphodiester linkage at a nick in DNA. In Step 1, the -amino group of a lysine residue of DNA ligase attacks the phosphorus atom bonded to the 5'-oxygen atom of the adenosine moiety of NAD+. Nicotinamide mononucleotide (NMN+) is displaced, generating an AMP–DNA-ligase intermediate. (With DNA ligases that use ATP as the cosubstrate, pyrophosphate is displaced.) In Step 2, an oxygen atom of the free 5'-phosphate group of the DNA attacks the phosphate group of the AMP-enzyme complex, forming an ADP-DNA intermediate. In Step 3, the nucleophilic 3'-hydroxyl group on the terminal residue of the adjacent DNA strand attacks the activated group of ADPDNA, releasing AMP and generating a phosphodiester linkage that seals the nick in the DNA strand. B represents any base.
 is displaced, generating an AMP–DNA-ligase intermediate. (With DNA ligases that use ATP as the cosubstrate, pyrophosphate is displaced.) In Step 2, an oxygen atom of the free 5 -phosphate group of the DNA attacks the phosphate group of the AMP-enzyme complex, forming an ADP-DNA intermediate. In Step 3, the nucleophilic 3 -hydroxyl group on the terminal residue of the adjacent DNA strand attacks the activated group of ADPDNA, releasing AMP and generating a phosphodiester linkage that seals the nick in the DNA strand. B represents any base.")
40
Figure 20.14 Proposed mechanism of DNA ligase in E. coli. Using NAD+ as a cosubstrate, E. coli DNA ligase catalyzes the formation of a phosphodiester linkage at a nick in DNA. In Step 1, the -amino group of a lysine residue of DNA ligase attacks the phosphorus atom bonded to the 5'-oxygen atom of the adenosine moiety of NAD+. Nicotinamide mononucleotide (NMN+) is displaced, generating an AMP–DNA-ligase intermediate. (With DNA ligases that use ATP as the cosubstrate, pyrophosphate is displaced.) In Step 2, an oxygen atom of the free 5'-phosphate group of the DNA attacks the phosphate group of the AMP-enzyme complex, forming an ADP-DNA intermediate. In Step 3, the nucleophilic 3'-hydroxyl group on the terminal residue of the adjacent DNA strand attacks the activated group of ADPDNA, releasing AMP and generating a phosphodiester linkage that seals the nick in the DNA strand. B represents any base.
 is displaced, generating an AMP–DNA-ligase intermediate. (With DNA ligases that use ATP as the cosubstrate, pyrophosphate is displaced.) In Step 2, an oxygen atom of the free 5 -phosphate group of the DNA attacks the phosphate group of the AMP-enzyme complex, forming an ADP-DNA intermediate. In Step 3, the nucleophilic 3 -hydroxyl group on the terminal residue of the adjacent DNA strand attacks the activated group of ADPDNA, releasing AMP and generating a phosphodiester linkage that seals the nick in the DNA strand. B represents any base.")
41
Figure 20.14 Proposed mechanism of DNA ligase in E. coli. Using NAD+ as a cosubstrate, E. coli DNA ligase catalyzes the formation of a phosphodiester linkage at a nick in DNA. In Step 1, the -amino group of a lysine residue of DNA ligase attacks the phosphorus atom bonded to the 5'-oxygen atom of the adenosine moiety of NAD+. Nicotinamide mononucleotide (NMN+) is displaced, generating an AMP–DNA-ligase intermediate. (With DNA ligases that use ATP as the cosubstrate, pyrophosphate is displaced.) In Step 2, an oxygen atom of the free 5'-phosphate group of the DNA attacks the phosphate group of the AMP-enzyme complex, forming an ADP-DNA intermediate. In Step 3, the nucleophilic 3'-hydroxyl group on the terminal residue of the adjacent DNA strand attacks the activated group of ADPDNA, releasing AMP and generating a phosphodiester linkage that seals the nick in the DNA strand. B represents any base.
 is displaced, generating an AMP–DNA-ligase intermediate. (With DNA ligases that use ATP as the cosubstrate, pyrophosphate is displaced.) In Step 2, an oxygen atom of the free 5 -phosphate group of the DNA attacks the phosphate group of the AMP-enzyme complex, forming an ADP-DNA intermediate. In Step 3, the nucleophilic 3 -hydroxyl group on the terminal residue of the adjacent DNA strand attacks the activated group of ADPDNA, releasing AMP and generating a phosphodiester linkage that seals the nick in the DNA strand. B represents any base.")
42
20.4 Model of the Replisome Replisome contains: a primosome, DNA polymerase III holoenzyme, additional proteins DnaB helicase is part of the primosome and facilitates unwinding of the DNA helix Topoisomerases relieve supercoiling ahead of the replicating fork (not part of the replisome) Single-stranded binding proteins (SSBs) stabilize single-stranded DNA
 Single-stranded binding proteins (SSBs) stabilize single-stranded DNA.")
43
Fig 20.15 Replisome DNA synthesis

44
Fig (continued)
")
45
Fig (continued)
")
46
Fig (continued)
")
47
Animations DNA Elongation Leading and Lagging Strand Synthesis

48
20.5 Initiation and Termination of DNA Replication
Replisome assembles at origin site (oriC) Initial assembly depends on unwinding of the DNA caused by binding certain proteins DnaA is one initiation protein
 Initial assembly depends on unwinding of the DNA caused by binding certain proteins. DnaA is one initiation protein.")
49
Termination site (ter)
Terminator utilization substance (Tus) binds to the ter site Tus inhibits helicase activity and thus prevents replication forks continuing through this region
 binds to the ter site. Tus inhibits helicase activity and thus prevents replication forks continuing through this region.")
50
Fig 20. 16 Origin (oriC) and terminus (ter) of DNA replication in E
Fig Origin (oriC) and terminus (ter) of DNA replication in E. coli dnaA is gene for protein DnaA (required to initiate replication) Red arrows indicate direction of movement of replication forks
 and terminus (ter) of DNA replication in E. coli. dnaA is gene for protein DnaA (required to initiate replication) Red arrows indicate direction of movement of replication forks.")
51
Fig 20.17 E. coli Tus bound to DNA
Tus binds to specific sequences at the termination site of DNA replication Tus blocks movement of the replisome by inhibiting the helicase activity of the replisome.

52
Box 20.1 Sequencing DNA Using Dideoxynucleotides
Sanger method uses 2’,3’-dideoxynucleoside triphosphates (ddNTPs) which are incorporated at the 3’ end of a growing chain in place of a dNTP Since ddNTPs lack a 3’-hydroxyl group, subsequent nucleotide addition cannot take place Small amounts of ddNTP’s terminate replication of some chains at each step, leaving a set of fragments of different lengths
 which are incorporated at the 3’ end of a growing chain in place of a dNTP. Since ddNTPs lack a 3’-hydroxyl group, subsequent nucleotide addition cannot take place. Small amounts of ddNTP’s terminate replication of some chains at each step, leaving a set of fragments of different lengths.")
53
Chemical structure of a ddNTP

54
Box 20.1 Sanger method for sequencing DNA

55
Sanger method for sequencing DNA

56
Sanger method (continued)
")
57
Sanger method (continued)
")
58
Automated Dideoxy DNA Sequencing with Fluorescent ddNTPs
(

59
20.6 DNA Replication in Eukaryotes
Mechanism similar to that in prokaryotes: leading strand continuous synthesis, lagging strand discontinuous synthesis Replication forks move more slowly, but many replication forks Okazaki fragments are shorter in eukaryotes (~ residues) At least 5 different DNA polymerases
 At least 5 different DNA polymerases.")
60
Fig 20.18 Eukaryotic cell division cycle

61
Table 20.2

62
Accessory Proteins Associated with the Replication Fork
PCNA (proliferating cell nuclear antigen) forms structure resembling b-subunit sliding clamp (E. coli DNA polymerase III) RPC (replication factor C) similar to g complex of DNA pol III RPA (replication factor A) similar to prokaryotic SSB Helicases also present to unwind DNA
 forms structure resembling b-subunit sliding clamp (E. coli DNA polymerase III) RPC (replication factor C) similar to g complex of DNA pol III. RPA (replication factor A) similar to prokaryotic SSB. Helicases also present to unwind DNA.")
63
Animation DNA Replication

64
20.7 Repair of Damaged DNA DNA is the only cellular macromolecule that can be repaired DNA damage includes: base modifications nucleotide deletions or insertions cross-linking of DNA strands breakage of phosphodiester backbone

65
DNA Repair Mechanisms Specific repair enzymes scan DNA to detect any alterations Lesions may be fixed by direct repair, which does not require breaking the phosphodiester backbone of DNA Repair protects individual cells and subsequent generations

66
A. Repair after Photodimerization: An Example of Direct Repair
Double-helical DNA is very sensitive to damage by ultaviolet (UV) light Dimerization of adjacent pyrimidines in a DNA strand is common (e.g. thymines) Replication cannot proceed in the presence of pyrimidine dimers (distort the template strand) Thymine dimers are repaired in all organisms
 light. Dimerization of adjacent pyrimidines in a DNA strand is common (e.g. thymines) Replication cannot proceed in the presence of pyrimidine dimers (distort the template strand) Thymine dimers are repaired in all organisms.")
67
Fig 20.19 Photodimerization of adjacent thymines

68
Fig 20.20 Repair of thymine dimers by DNA photolyase

69
Fig 20.20

70
Fig (cont)
")
71
Fig (cont)
")
72
B. Excision Repair DNA can be damaged by alkylation, methylation, deamination, loss of heterocyclic bases (depurination or depyrimidization) General excision-repair pathway can repair many of these defects Overall pathway is similar in all organisms

73
Fig 20.21 General excision-repair pathway

74
Fig 20.21

75
Fig (cont)
")
76
Fig 20.22 Hydrolytic deamination of cytosine to uracil
Uracil in place of cytosine causes incorporation of an incorrect base during replication DNA glycosylases hydrolyze base-sugar N-glycosidic bonds Deaminated bases are then removed and replaced

77
Fig 20.23 Uracil N-glycosylase (human mitochondria)
Enzyme is bound to a uracil-containing nucleotide (green) that has been flipped out of the stacked region of DNA
 that has been flipped out of the stacked region of DNA.")
78
Fig 20.24 Repair of damage from deamination of cytosine

79
(AP site: apurine and apyrimidine sites)
Fig 20.24 (AP site: apurine and apyrimidine sites)
")
80
Fig (cont)
")
81
Fig (cont)
")
82
20.8 Homologous Recombination
Recombination - exchange or transfer of pieces of DNA from one chromosome to another or within a chromosome Homologous recombination - occurs between pieces of DNA that have closely related sequences Nonhomologous recombination occurs between unrelated sequences (e.g. Transposons )
")
83
A. The Holliday Model of General Recombination
Figure 20.25

84
Figure 20.25

85
Fig (cont)
")
86
B. Recombination in E. coli
Recombination starts with generation of single-stranded DNA with a free 3’ end In E. coli, RecBCD endonuclease binds to DNA, cleaves one strand, then unwinds DNA in an ATP-dependent reaction Strand exchange begins when single-stranded DNA invades a neighboring double helix Rec A is a strand exchange protein

87
Fig 20.26 Strand exchange catalyzed by RecA

88
Fig 20.26 Figure 20.26 Strand exchange catalyzed by RecA.

89
Fig (cont) Figure 20.26 Strand exchange catalyzed by RecA.
 Figure Strand exchange catalyzed by RecA.")
90
C. Recombination Can Be a Form of Repair
Recombination enzymes probably evolved to help DNA repair (confers a selective advantage) Recombination also creates new combinations of genes on a chromosome, increasing chances of evolutionary survival
 Recombination also creates new combinations of genes on a chromosome, increasing chances of evolutionary survival.")
91
Fig 20.27 Action of Ruv proteins at Holliday junctions

92
Fig 20.28 Model of RuvA and RuvB bound to Holliday junction

93
Fig 20.29 Branch migration and resolution

94
Box 20.2 Molecular Links Between DNA Repair and Breast Cancer
Some cancers are due to mutations in the genes BRCA1 or BRCA2 BRCA1 and BRCA2 are required for normal recombinational repair of double strand breaks (DSB) Ionizing radiation induces nuclear foci of the DNA repair protein BRCA1. Energetic can induce double-stranded breaks in DNA and trigger DNA repair. This tissue culture cell nucleus was exposed to IR and then treated with antibodies that recognize BRCA1 (stained green).
 Ionizing radiation induces nuclear foci of the DNA repair protein BRCA1. Energetic can induce double-stranded breaks in DNA and trigger DNA repair. This tissue culture cell nucleus was exposed to IR and then treated with antibodies that recognize BRCA1 (stained green).")
96
Fig 20.20 PCNA (proliferating cell nuclear antigen)
Trimeric protein that forms sliding clamp that surrounds DNA Part of eukaryotic replisome

97
Fig 20.28 Stereo view of a Holliday junction structure










 Accurate Need to correct replication errors and repair damage.>")


 Required reading: Stryer’s Biochemistry 5 th edition p. 127-128, 750-754, 759-766, 768-773 (or Stryer’s Biochemistry.>")










