Download presentation
Presentation is loading. Please wait.

1
Non-coding RNAs (ncRNAs)
ncRNAs: A brief intro Long non-coding RNAs (lncRNAs or lincRNAs): >200 bp miRNAs: Biogenesis, measurement, functional analysis, & utility Key references: Prensner JR & Chinnaiyan AM. The emergence of lincRNAs in cancer biology. Cancer Disc. 1: , 2011. 2. Guttman M and Rinn JL. Modular regulatory principles of large non-coding RNAs. Nature 482, , 2012. 3. Lee JT. Epigenetic regulation by long noncoding RNAs. Science 338: , 2012. Dean G. Tang (Molecular Biology of Cancer; 3/6/2013) Acknowledgement: Julie Liu (Tang lab); David Brown (MiRna Therapeutics); Thomson/Hammond (UNC)
: >200 bp. miRNAs: Biogenesis, measurement, functional analysis, & utility. Key references: Prensner JR & Chinnaiyan AM. The emergence of lincRNAs in cancer biology. Cancer Disc. 1: , Guttman M and Rinn JL. Modular regulatory principles of large non-coding RNAs. Nature 482, , Lee JT. Epigenetic regulation by long noncoding RNAs. Science 338: , Dean G. Tang (Molecular Biology of Cancer; 3/6/2013) Acknowledgement: Julie Liu (Tang lab); David Brown (MiRna Therapeutics); Thomson/Hammond (UNC)")
2
Encylcopedia of DNA Elements (EnCODE)/Sept 2012
Science 337: , 2012 (32 groups; 24 exp/120 TF)) 76% of the genome is transcribed; 18,400 (8,800 sRNA+9,600 lncRNA); 2.89 million DnaseI sites (1/3 in each cell type and 3,700 in all cells); >3.9 million TF binding sites >70,000 ‘promoter’ regions ~400,000 ‘enhancer’ regions 11,224 pseudogenes 15 terabites of data 5C: Chromosome Conformation Capture Carbon Copy. Detection of chromosome interactions. Physical interactions between genetic elements located throughout the genome play important roles in gene regulation and can be identified with the Chromosome Conformation Capture (3C) methodology. 3C converts physical chromatin interactions into specific ligation products, which are quantified individually by PCR. Here we present a high-throughput 3C approach, 3C-Carbon Copy (5C), that employs microarrays or quantitative DNA sequencing using 454-technology as detection methods. We applied 5C to analyze a 400-kb region containing the human β-globin locus and a 100-kb conserved gene desert region. We validated 5C by detection of several previously identified looping interactions in the β-globin locus. We also identified a new looping interaction in K562 cells between the β-globin Locus Control Region and the γ–β-globin intergenic region. Interestingly, this region has been implicated in the control of developmental globin gene switching. 5C should be widely applicable for large-scale mapping of cis- and trans- interaction networks of genomic elements and for the study of higher-order chromosome structure (Dostie J et al., Genome Res. 16: , 2006). FAIRE-Seq (Formaldehyde-Assisted Isolation of Regulatory Elements) is a method in molecular biology used for determining the sequences of those DNA regions in the genome associated with regulatory activity.[1] The technique was developed in the laboratory of Jason D. Lieb at the University of North Carolina, Chapel Hill. In contrast to DNase-Seq, the FAIRE-Seq protocol doesn't require the permeabilization of cells or isolation of nuclei, and can analyse any cell types. In a study of seven diverse human cell types, DNase-seq and FAIRE-seq produced strong cross-validation, with each cell type having 1-2% of the human genome as open chromatin. The two methods are not fully overlapping, with FAIRE being more sensitive to find distal regulatory elements that are not detected with DNase-Seq but missing promoter regions that are detected with DNase-Seq. [2][3] The protocol is based on the fact that the formaldehyde cross-linking is more efficient in nucleosome-bound DNA than it is in nucleosome-depleted regions of the genome. This method then segregates the non cross-linked DNA that is usually found in open chromatin, which is then sequenced. The protocol consists of cross linking, phenol extraction and sequencing the DNA in aqueous phase. *Bioinformatic analysis of the chromatin marks in intergenic DNA regions and of ESTs predicts >5,000 lncRNA genes in the human genome (Guttman et al., Nature 458, , 2009; Khalil AM et al., PNAS 106, , 2009). *Including antisense, intronic, intergenic, pseudogenes and retrotransposons. *The most ‘famous’ lincRNAs include Xist, H19, Air, Hotair, etc, which all seem to operate at the transcriptional level by binding to proteins in histone-modifying complexes and targeting them to particular genes.
) 76% of the genome is transcribed; 18,400 (8,800 sRNA+9,600 lncRNA); 2.89 million DnaseI sites (1/3 in each. cell type and 3,700 in all cells); >3.9 million TF binding sites. >70,000 ‘promoter’ regions. ~400,000 ‘enhancer’ regions. 11,224 pseudogenes. 15 terabites of data. 5C: Chromosome Conformation Capture Carbon Copy. Detection of chromosome interactions. Physical interactions between genetic elements located throughout the genome play important roles in gene regulation and can be identified with the Chromosome Conformation Capture (3C) methodology. 3C converts physical chromatin interactions into specific ligation products, which are quantified individually by PCR. Here we present a high-throughput 3C approach, 3C-Carbon Copy (5C), that employs microarrays or quantitative DNA sequencing using 454-technology as detection methods. We applied 5C to analyze a 400-kb region containing the human β-globin locus and a 100-kb conserved gene desert region. We validated 5C by detection of several previously identified looping interactions in the β-globin locus. We also identified a new looping interaction in K562 cells between the β-globin Locus Control Region and the γ–β-globin intergenic region. Interestingly, this region has been implicated in the control of developmental globin gene switching. 5C should be widely applicable for large-scale mapping of cis- and trans- interaction networks of genomic elements and for the study of higher-order chromosome structure (Dostie J et al., Genome Res. 16: , 2006). FAIRE-Seq (Formaldehyde-Assisted Isolation of Regulatory Elements) is a method in molecular biology used for determining the sequences of those DNA regions in the genome associated with regulatory activity.[1] The technique was developed in the laboratory of Jason D. Lieb at the University of North Carolina, Chapel Hill. In contrast to DNase-Seq, the FAIRE-Seq protocol doesn t require the permeabilization of cells or isolation of nuclei, and can analyse any cell types. In a study of seven diverse human cell types, DNase-seq and FAIRE-seq produced strong cross-validation, with each cell type having 1-2% of the human genome as open chromatin. The two methods are not fully overlapping, with FAIRE being more sensitive to find distal regulatory elements that are not detected with DNase-Seq but missing promoter regions that are detected with DNase-Seq. [2][3] The protocol is based on the fact that the formaldehyde cross-linking is more efficient in nucleosome-bound DNA than it is in nucleosome-depleted regions of the genome. This method then segregates the non cross-linked DNA that is usually found in open chromatin, which is then sequenced. The protocol consists of cross linking, phenol extraction and sequencing the DNA in aqueous phase. *Bioinformatic analysis of the chromatin marks in intergenic DNA regions and of ESTs predicts >5,000 lncRNA genes. in the human genome (Guttman et al., Nature 458, , 2009; Khalil AM et al., PNAS 106, , 2009). *Including antisense, intronic, intergenic, pseudogenes and retrotransposons. *The most ‘famous’ lincRNAs include Xist, H19, Air, Hotair, etc, which all seem to operate at the transcriptional. level by binding to proteins in histone-modifying complexes and targeting them to particular genes.")
3
Types of ncRNAs Prensner JR & Chinnaiyan AM. Cancer Disc. 1: , 2011.

4
Critical features of lncRNAs

5
Examples of lncRNAs Prensner JR & Chinnaiyan AM. Cancer Disc. 1: , 2011.

6
Gene expression regulation by lncRNAs Prensner JR & Chinnaiyan AM.
Cancer Disc. 1: , 2011.

7
Mechanisms of X-inactivation
In mammals, XCI is triggered by Xist RNA to equalize gene expression between the sexes. The random form of XCI occurs ONLY once on E , when the epiblast has cells. Beyond E5.5, the inactive X (Xi; the Barr body) enters into a ‘maintenance phase’ in which the same X chromosome is propagated as Xi for the remainder of female life. The X-inactivation center (XIC), ~100kb, contains several noncoding RNA genes Xist, Tsix, Jpx/Enox, and Xite. There are also 2 protein-coding genes (Tsx and Cnbp2). The Xist promoter is the master regulator of X inactivation. Initiation of XCI depends on Xist (the 17 kb X-inactive specific transcript) that targets and tethers PRCs to the X chromosome in cis. Xist is dispensable once the Xi is established. Xi Xist expression Tsix expression Xa Xist expression Tsix expression Chromosome-wide silencing requires Xist RNA, which spreads along the X and presumably recruits silencing factors to that chromosome. Xist is regulated in cis by its antisense partner, Tsix, which is responsible for keeping the X-chromosome inactive. Tsix is in turn regulated by Xite, an upstream locus harboring multiple intergenic transcripts and a Tsix-specific enhancer.
 enters into a ‘maintenance phase’ in which the same X chromosome is propagated as Xi for the remainder of female life. The X-inactivation center (XIC), ~100kb, contains several noncoding RNA genes Xist, Tsix, Jpx/Enox, and Xite. There are also 2 protein-coding genes (Tsx and Cnbp2). The Xist promoter is the master regulator of X inactivation. Initiation of XCI depends on Xist (the 17 kb X-inactive specific transcript) that targets and tethers PRCs to the X chromosome in cis. Xist is dispensable once the Xi is established. Xi Xist expression. Tsix expression. Xa Xist expression. Tsix expression. Chromosome-wide silencing requires Xist RNA, which spreads along the X and presumably recruits silencing factors to that chromosome. Xist is regulated in cis by its antisense partner, Tsix, which is responsible for keeping the X-chromosome inactive. Tsix is in turn regulated by Xite, an upstream locus harboring multiple intergenic transcripts and a Tsix-specific enhancer.")
8
Mechanisms of X-inactivation
Lee JT, Science 2012

9
Mechanisms of X-inactivation
PRC2, H3K27me3 RepA binding to EZH2 Tsix acting as decoy Conditional deletion of Xist in blood cells: Born alive, viable, but females die around 2 months Massive spleen in female, hyperplasia (early stage) to leukemia (late stage). Progressive bone marrow disease, myelofibrosis, leukemia (mixed MPN/MDS), and histiocytic sarcoma. (Yildirim E et al., Xist RNA is a potent suppressor of hematologic cancer in mice. Cell 152, , 2013).
 to leukemia (late stage). Progressive bone marrow disease, myelofibrosis, leukemia (mixed MPN/MDS), and histiocytic sarcoma. (Yildirim E et al., Xist RNA is a potent suppressor of hematologic cancer in mice. Cell 152, , 2013).")
10
Genome regulation by long noncoding RNA (H. Chang)
HOTAIR (HOX antisense intergenic RNA): on chr.12, encodes a 2.2kb lncRNA. HOTAIR is located in the HoxC cluster, interacts with Suz12, EZH2, and LSD1 as a scaffold, and silences HoxD cluster. HOTAIR is upregulated in many cancers (br cancer and HCC) HOTTIP & HOTAIRM1: located at opposite ends of the HOXA locus, activating HOXA transcription. NEST: chr.12 Controls susceptibility of virus infection; Interacts with WRD5, promotes H3K4me3 at INFG, and activates INFG. overexpressionresistant for pathogen challenge. first known lincRNA that can regulate the chromatin state of genes on distantly located chromosomes Gomez JA et al., Cell 152: , 2013.
: on chr.12, encodes a 2.2kb lncRNA. HOTAIR is located in the HoxC cluster, interacts with Suz12, EZH2, and LSD1 as a scaffold, and silences HoxD cluster. HOTAIR is upregulated in many cancers (br cancer and HCC) HOTTIP & HOTAIRM1: located at opposite ends of the HOXA locus, activating HOXA transcription. NEST: chr.12. Controls susceptibility of virus infection; Interacts with WRD5, promotes H3K4me3 at INFG, and activates INFG. overexpressionresistant for pathogen challenge. first known lincRNA that can regulate the chromatin state of genes on distantly located chromosomes. Gomez JA et al., Cell 152: ,")
11
lncRNAs in prostate cancer (A. Chinnaiyan)
HOTAIR: highly expressed in breast Ca, lung Ca, but low in PCa. PCAT-1, a novel prostate-specific regulator of cell proliferation, a transcriptional repressor, and a target of PRC2. It’s a 1.9 kb pA-containing lncRNA comprised of 2 exons and located in the Chr8q24 gene desert (Presener JR et al., Nature Biotech 29:742-9, 2011). SChLAP1 (Second Chromosome Locus Associated with Prostate-1), a >500kb locus in a gene desert in Chr2q31, including PCAT-109, PCAT-114. SChLAP1 over-expressed in 20% PCa patients. Correlates with poor outcome, more aggressive samples.
. SChLAP1 (Second Chromosome Locus Associated with Prostate-1), a >500kb locus in a gene desert in Chr2q31, including PCAT-109, PCAT-114. SChLAP1 over-expressed in 20% PCa patients. Correlates with poor outcome, more aggressive samples.")
12
Biological functions:
SChLAP1 expression: Nuclear staining in VCaP, 22Rv1, LNCaP. In situ hybridization of FFPE PCa revealed high expression levels. Biological functions: Promotes invasion, (not strong phenotype in proliferation) KD with shRNA in vitro (in cells) and in vivo (cardiac injection) results decrease in invasion and met. spread. Molecular mechanisms: Microarray of gene expression of a serial of PCa KD of SchLAP1. Reversed relationship with SWN/SNF complex. Pull down SNF5, also pull down SChLAP1. ChIP –seq of SNF5 in RWPE-SchLAP1 OE shows reduced binding.
 KD with shRNA in vitro (in cells) and in vivo (cardiac injection) results decrease in invasion and met. spread. Molecular mechanisms: Microarray of gene expression of a serial of PCa KD of SchLAP1. Reversed relationship with SWN/SNF complex. Pull down SNF5, also pull down SChLAP1. ChIP –seq of SNF5 in RWPE-SchLAP1 OE shows reduced binding.")
13
Mechanisms of lncRNA function
Prensner JR & Chinnaiyan AM. Cancer Disc. 1: , 2011.

14
Mechanisms of lncRNA function
Lee JT, Science 2012

15
Mechanisms of lncRNA function
Lee JT, Science 2012

16
Non-coding RNAs (ncRNAs)
ncRNAs: A brief intro Long non-coding RNAs (lncRNAs or lincRNAs): >200 bp miRNAs: Biogenesis, measurement, functional analysis, utility Key references: Prensner JR & Chinnaiyan AM. The emergence of lincRNAs in cancer biology. Cancer Disc. 1: , 2011. 2. Guttman M and Rinn JL. Modular regulatory principles of large non-coding RNAs. Nature 482, , 2012. 3. Lee JT. Epigenetic regulation by long noncoding RNAs. Science 338: , 2012. Dean G. Tang (Molecular Biology of Cancer; 3/6/2013)
: >200 bp. miRNAs: Biogenesis, measurement, functional analysis, utility. Key references: Prensner JR & Chinnaiyan AM. The emergence of lincRNAs in cancer biology. Cancer Disc. 1: , Guttman M and Rinn JL. Modular regulatory principles of large non-coding RNAs. Nature 482, , Lee JT. Epigenetic regulation by long noncoding RNAs. Science 338: , Dean G. Tang (Molecular Biology of Cancer; 3/6/2013)")
18
Exonic Non-coding Intronic Non-coding and coding
MicroRNAs are transcribed by RNA Polymerase II Thomson, J. M/Hammond S Exonic Non-coding Intronic Non-coding and coding Lee, et al., EMBO J. 23:

19
Thomson, J. M/Hammond S

20
Pasquinelli, Nat Rev Genetics, 2012 Vol 13 No 4

21
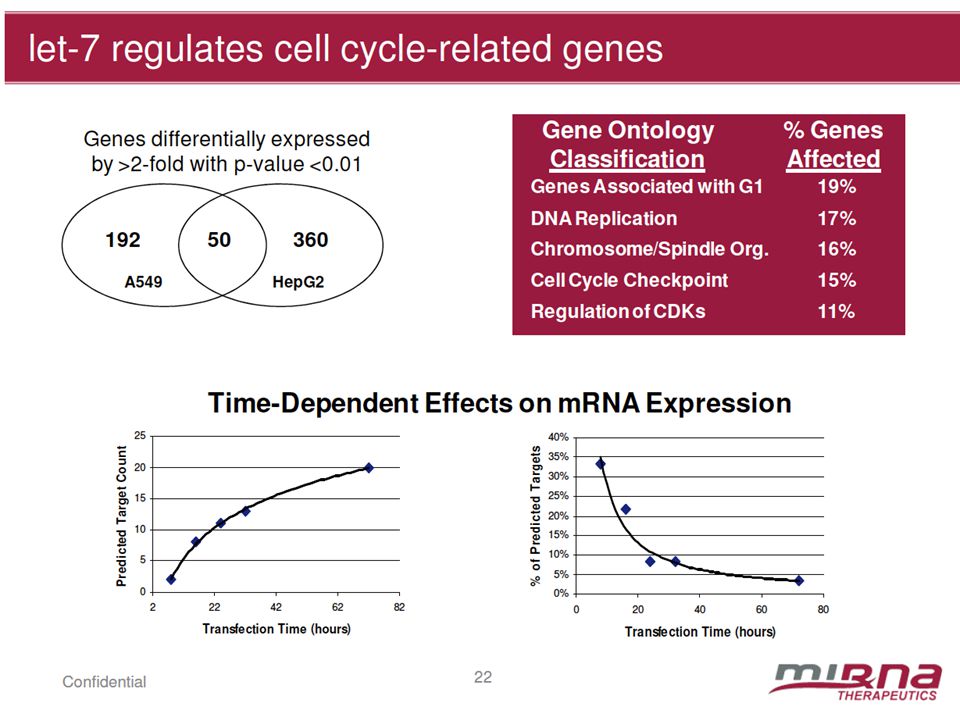
Nijiro Nohata, et al. ELSEVIER.2012

22
*For microprocessor recognition, sequences
within 40 nt upstream and 40 nt downstream of the pre-miRNA hairpins are required. *Most C. elegans pri-miRNAs lack determinants required for processing in human cells. *Pairing in the basal stem is important. *Primary sequence features, including the basal UG, the CNNC, and the apical GUG motifs, contribute to efficient processing in human cells. *79% of the conserved human miRNAs contain at least one of the three motifs. *These motifs are not enriched in C. elegans pri- miRNAs and, when added to the C. elegans pri- miRNAs, confer more efficient processing in mammalian cells. Auyeung VC, et al. Cell 152: , 2013

23
Krol, Loedige &Filipowicz, Nat Rev Genetics, 2010 Vol 11 No 9
MicroRNAs (miRNAs) are processed from RNA polymerase II (RNAPII)-specific transcripts of independent genes or from introns of protein-coding genes2, 5. In the canonical pathway, primary precursor (pri-miRNA) processing occurs in two steps, catalysed by two members of the RNase III family of enzymes, Drosha and Dicer, operating in complexes with dsRNA-binding proteins (dsRBPs), for example DGCR8 and transactivation-responsive (TAR) RNA-binding protein (TRBP) in mammals. In the first nuclear step, the Drosha–DGCR8 complex processes pri-miRNA into an ~70-nucleotide precursor hairpin (pre-miRNA), which is exported to the cytoplasm. Some pre-miRNAs are produced from very short introns (mirtrons) as a result of splicing and debranching, thereby bypassing the Drosha–DGCR8 step. In either case, cleavage by Dicer, assisted by TRBP, in the cytoplasm yields an ~20-bp miRNA/miRNA* duplex. In mammals, argonaute 2 (AGO2), which has robust RNaseH-like endonuclease activity, can support Dicer processing by cleaving the 3′ arm of some pre-miRNAs, thus forming an additional processing intermediate called AGO2-cleaved precursor miRNA (ac-pre-miRNA)70. Processing of pre-miR-451 also requires cleavage by AGO2, but is independent of Dicer and the 3′ end is generated by exonucleolytic trimming66, 67. Following processing, one strand of the miRNA/miRNA* duplex (the guide strand) is preferentially incorporated into an miRNA-induced silencing complex (miRISC), whereas the other strand (passenger or miRNA*) is released and degraded (not shown). Generally, the retained strand is the one that has the less stably base-paired 5′ end in the miRNA/miRNA* duplex. miRNA* strands are not always by-products of miRNA biogenesis and can also be loaded into miRISC to function as miRNAs26, 58, 59, 60. See Box 2 for details of miRISC function. GW182, glycine-tryptophan protein of 182 kDa; m7G, 7-methylguanosine-cap; PABP, poly(A) binding protein. Krol, Loedige &Filipowicz, Nat Rev Genetics, 2010 Vol 11 No 9
 are processed from RNA polymerase II (RNAPII)-specific transcripts of independent genes or from introns of protein-coding genes2, 5. In the canonical pathway, primary precursor (pri-miRNA) processing occurs in two steps, catalysed by two members of the RNase III family of enzymes, Drosha and Dicer, operating in complexes with dsRNA-binding proteins (dsRBPs), for example DGCR8 and transactivation-responsive (TAR) RNA-binding protein (TRBP) in mammals. In the first nuclear step, the Drosha–DGCR8 complex processes pri-miRNA into an ~70-nucleotide precursor hairpin (pre-miRNA), which is exported to the cytoplasm. Some pre-miRNAs are produced from very short introns (mirtrons) as a result of splicing and debranching, thereby bypassing the Drosha–DGCR8 step. In either case, cleavage by Dicer, assisted by TRBP, in the cytoplasm yields an ~20-bp miRNA/miRNA* duplex. In mammals, argonaute 2 (AGO2), which has robust RNaseH-like endonuclease activity, can support Dicer processing by cleaving the 3′ arm of some pre-miRNAs, thus forming an additional processing intermediate called AGO2-cleaved precursor miRNA (ac-pre-miRNA)70. Processing of pre-miR-451 also requires cleavage by AGO2, but is independent of Dicer and the 3′ end is generated by exonucleolytic trimming66, 67. Following processing, one strand of the miRNA/miRNA* duplex (the guide strand) is preferentially incorporated into an miRNA-induced silencing complex (miRISC), whereas the other strand (passenger or miRNA*) is released and degraded (not shown). Generally, the retained strand is the one that has the less stably base-paired 5′ end in the miRNA/miRNA* duplex. miRNA* strands are not always by-products of miRNA biogenesis and can also be loaded into miRISC to function as miRNAs26, 58, 59, 60. See Box 2 for details of miRISC function. GW182, glycine-tryptophan protein of 182 kDa; m7G, 7-methylguanosine-cap; PABP, poly(A) binding protein. Krol, Loedige &Filipowicz, Nat Rev Genetics, 2010 Vol 11 No 9.")
24
David & McCray Jr, Nat Rev Genetics, 2011 Vol 12 No 5
Primary microRNAs (pri-miRNAs) are transcribed by RNA polymerases156, 157, 158 and are trimmed by the microprocessor complex (comprising Drosha and microprocessor complex subunit DCGR8) into ~70 nucleotide precursors, called pre-miRNAs67, 159, 160 (left side of the figure). miRNAs can also be processed from spliced short introns (known as mirtrons)161. pre-miRNAs contain a loop and usually have interspersed mismatches along the duplex. pre-miRNAs associate with exportin 5 and are exported to the cytoplasm162, 163, where a complex that contains Dicer, TAR RNA-binding protein (TRBP; also known as TARBP2) and PACT (also known as PRKRA) processes the pre-miRNAs into miRNA–miRNA* duplexes116, 164, 165. The duplex associates with an Argonaute (AGO) protein within the precursor RNAi-induced silencing complex (pre-RISC). One strand of the duplex (the passenger strand) is removed. The mature RISC contains the guide strand, which directs the complex to the target mRNA for post-transcriptional gene silencing. The 'seed' region of an miRNA is indicated; in RNAi trigger design, the off-target potential of this sequence needs to be considered. Long dsRNAs (right side of the figure) are processed by Dicer, TRBP and PACT into small interfering RNAs (siRNAs). siRNAs are 20–24-mer RNAs and harbour 3′OH and 5′ phosphate (PO4) groups, with 3′ dinucleotide overhangs3, 166, 167. Within the pre-RISC complex, an AGO protein cleaves the passenger siRNA strand. Then, the mature RISC, containing an AGO protein and the guide strand, associates with the target mRNA for cleavage. The inset shows the properties of siRNAs. The thermodynamic stability of the terminal sequences will direct strand loading. Like naturally occurring or artificially engineered miRNAs, the potential 'seed' region can be a source for miRNA-like off-target silencing. shRNA, short hairpin RNA.
 are transcribed by RNA polymerases156, 157, 158 and are trimmed by the microprocessor complex (comprising Drosha and microprocessor complex subunit DCGR8) into ~70 nucleotide precursors, called pre-miRNAs67, 159, 160 (left side of the figure). miRNAs can also be processed from spliced short introns (known as mirtrons)161. pre-miRNAs contain a loop and usually have interspersed mismatches along the duplex. pre-miRNAs associate with exportin 5 and are exported to the cytoplasm162, 163, where a complex that contains Dicer, TAR RNA-binding protein (TRBP; also known as TARBP2) and PACT (also known as PRKRA) processes the pre-miRNAs into miRNA–miRNA* duplexes116, 164, 165. The duplex associates with an Argonaute (AGO) protein within the precursor RNAi-induced silencing complex (pre-RISC). One strand of the duplex (the passenger strand) is removed. The mature RISC contains the guide strand, which directs the complex to the target mRNA for post-transcriptional gene silencing. The seed region of an miRNA is indicated; in RNAi trigger design, the off-target potential of this sequence needs to be considered. Long dsRNAs (right side of the figure) are processed by Dicer, TRBP and PACT into small interfering RNAs (siRNAs). siRNAs are 20–24-mer RNAs and harbour 3′OH and 5′ phosphate (PO4) groups, with 3′ dinucleotide overhangs3, 166, 167. Within the pre-RISC complex, an AGO protein cleaves the passenger siRNA strand. Then, the mature RISC, containing an AGO protein and the guide strand, associates with the target mRNA for cleavage. The inset shows the properties of siRNAs. The thermodynamic stability of the terminal sequences will direct strand loading. Like naturally occurring or artificially engineered miRNAs, the potential seed region can be a source for miRNA-like off-target silencing. shRNA, short hairpin RNA.")
25
Mechanisms of gene regulation by miRNAs
MicroRNAs (miRNAs) regulate gene expression through multiple pathways7. A complex of eukaryotic initiation factors (eIFs) binds the 5′ cap and the cytoplasmic poly(A)-binding protein (PABPC), connecting the 5′ and 3′ ends of mRNAs and stimulating their translation111 by the ribosome (shown in pink). a | Perfect pairing between an miRNA and its target site induces endonucleolytic cleavage by Argonaute (AGO), leading to rapid degradation of the mRNA. b | Partial pairing of the miRNA complex to target 3′ untranslated region (UTR) sites can result in deadenylation of the mRNA through recruitment of the CCR4–NOT complex by the miRNA-induced silencing complex (miRISC)-associated GW182 proteins. Loss of the poly(A) tail causes dissociation of PABPC and leads to degradation of the mRNA. c | The miRISC can also induce translational repression by blocking initiation via recruitment of CCR4–NOT by GW182. d | Translational repression can also be induced by the miRISC by inhibiting a step after initiation, such as promoting ribosome drop-off or stimulating proteolysis of the nascent peptide. e | miRNAs have also been shown to upregulate target expression under certain conditions through a mechanism that involves Argonaute and fragile X mental retardation protein 1 (FMR1). Pasquinelli, Nat Rev Genetics, 2012 Vol 13 No 4
 regulate gene expression through multiple pathways7. A complex of eukaryotic initiation factors (eIFs) binds the 5′ cap and the cytoplasmic poly(A)-binding protein (PABPC), connecting the 5′ and 3′ ends of mRNAs and stimulating their translation111 by the ribosome (shown in pink). a | Perfect pairing between an miRNA and its target site induces endonucleolytic cleavage by Argonaute (AGO), leading to rapid degradation of the mRNA. b | Partial pairing of the miRNA complex to target 3′ untranslated region (UTR) sites can result in deadenylation of the mRNA through recruitment of the CCR4–NOT complex by the miRNA-induced silencing complex (miRISC)-associated GW182 proteins. Loss of the poly(A) tail causes dissociation of PABPC and leads to degradation of the mRNA. c | The miRISC can also induce translational repression by blocking initiation via recruitment of CCR4–NOT by GW182. d | Translational repression can also be induced by the miRISC by inhibiting a step after initiation, such as promoting ribosome drop-off or stimulating proteolysis of the nascent peptide. e | miRNAs have also been shown to upregulate target expression under certain conditions through a mechanism that involves Argonaute and fragile X mental retardation protein 1 (FMR1). Pasquinelli, Nat Rev Genetics, 2012 Vol 13 No 4.")
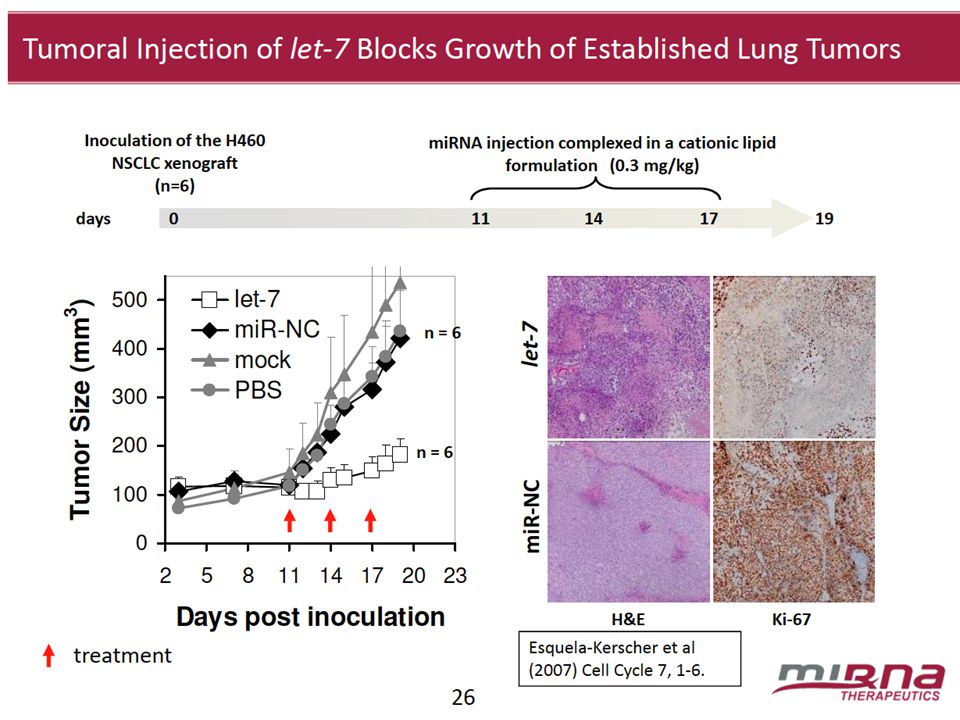
26
Evolutionary Conserved miRNA Cluster
Thomson, J. M/Hammond S

27
miR17-92 is Conserved in Vertebrates
Thomson, J. M/Hammond S miR17-92 is Conserved in Vertebrates

28
mRNAs as Regulators of miRNAs
ceRNA (competing endogenous RNA) MREs (microRNA response sequences) L Salmena,et al. Cell. 2011
 MREs (microRNA response sequences) L Salmena,et al. Cell")
29
Non-coding RNAs (ncRNAs)
ncRNAs: A brief intro Long non-coding RNAs (lncRNAs or lincRNAs): >200 bp miRNAs: Biogenesis, measurement, functional analysis, utility Key references: Prensner JR & Chinnaiyan AM. The emergence of lincRNAs in cancer biology. Cancer Disc. 1: , 2011. 2. Guttman M and Rinn JL. Modular regulatory principles of large non-coding RNAs. Nature 482, , 2012. 3. Lee JT. Epigenetic regulation by long noncoding RNAs. Science 338: , 2012. MicroRNAs (miRNAs) are processed from RNA polymerase II (RNAPII)-specific transcripts of independent genes or from introns of protein-coding genes2, 5. In the canonical pathway, primary precursor (pri-miRNA) processing occurs in two steps, catalysed by two members of the RNase III family of enzymes, Drosha and Dicer, operating in complexes with dsRNA-binding proteins (dsRBPs), for example DGCR8 and transactivation-responsive (TAR) RNA-binding protein (TRBP) in mammals. In the first nuclear step, the Drosha–DGCR8 complex processes pri-miRNA into an ~70-nucleotide precursor hairpin (pre-miRNA), which is exported to the cytoplasm. Some pre-miRNAs are produced from very short introns (mirtrons) as a result of splicing and debranching, thereby bypassing the Drosha–DGCR8 step. In either case, cleavage by Dicer, assisted by TRBP, in the cytoplasm yields an ~20-bp miRNA/miRNA* duplex. In mammals, argonaute 2 (AGO2), which has robust RNaseH-like endonuclease activity, can support Dicer processing by cleaving the 3′ arm of some pre-miRNAs, thus forming an additional processing intermediate called AGO2-cleaved precursor miRNA (ac-pre-miRNA)70. Processing of pre-miR-451 also requires cleavage by AGO2, but is independent of Dicer and the 3′ end is generated by exonucleolytic trimming66, 67. Following processing, one strand of the miRNA/miRNA* duplex (the guide strand) is preferentially incorporated into an miRNA-induced silencing complex (miRISC), whereas the other strand (passenger or miRNA*) is released and degraded (not shown). Generally, the retained strand is the one that has the less stably base-paired 5′ end in the miRNA/miRNA* duplex. miRNA* strands are not always by-products of miRNA biogenesis and can also be loaded into miRISC to function as miRNAs26, 58, 59, 60. See Box 2 for details of miRISC function. GW182, glycine-tryptophan protein of 182 kDa; m7G, 7-methylguanosine-cap; PABP, poly(A) binding protein. Dean G. Tang (Molecular Biology of Cancer; 3/6/2013)
: >200 bp. miRNAs: Biogenesis, measurement, functional analysis, utility. Key references: Prensner JR & Chinnaiyan AM. The emergence of lincRNAs in cancer biology. Cancer Disc. 1: , Guttman M and Rinn JL. Modular regulatory principles of large non-coding RNAs. Nature 482, , Lee JT. Epigenetic regulation by long noncoding RNAs. Science 338: , MicroRNAs (miRNAs) are processed from RNA polymerase II (RNAPII)-specific transcripts of independent genes or from introns of protein-coding genes2, 5. In the canonical pathway, primary precursor (pri-miRNA) processing occurs in two steps, catalysed by two members of the RNase III family of enzymes, Drosha and Dicer, operating in complexes with dsRNA-binding proteins (dsRBPs), for example DGCR8 and transactivation-responsive (TAR) RNA-binding protein (TRBP) in mammals. In the first nuclear step, the Drosha–DGCR8 complex processes pri-miRNA into an ~70-nucleotide precursor hairpin (pre-miRNA), which is exported to the cytoplasm. Some pre-miRNAs are produced from very short introns (mirtrons) as a result of splicing and debranching, thereby bypassing the Drosha–DGCR8 step. In either case, cleavage by Dicer, assisted by TRBP, in the cytoplasm yields an ~20-bp miRNA/miRNA* duplex. In mammals, argonaute 2 (AGO2), which has robust RNaseH-like endonuclease activity, can support Dicer processing by cleaving the 3′ arm of some pre-miRNAs, thus forming an additional processing intermediate called AGO2-cleaved precursor miRNA (ac-pre-miRNA)70. Processing of pre-miR-451 also requires cleavage by AGO2, but is independent of Dicer and the 3′ end is generated by exonucleolytic trimming66, 67. Following processing, one strand of the miRNA/miRNA* duplex (the guide strand) is preferentially incorporated into an miRNA-induced silencing complex (miRISC), whereas the other strand (passenger or miRNA*) is released and degraded (not shown). Generally, the retained strand is the one that has the less stably base-paired 5′ end in the miRNA/miRNA* duplex. miRNA* strands are not always by-products of miRNA biogenesis and can also be loaded into miRISC to function as miRNAs26, 58, 59, 60. See Box 2 for details of miRISC function. GW182, glycine-tryptophan protein of 182 kDa; m7G, 7-methylguanosine-cap; PABP, poly(A) binding protein. Dean G. Tang (Molecular Biology of Cancer; 3/6/2013)")
30
MicroRNAs (miRNAs) are processed from RNA polymerase II (RNAPII)-specific transcripts of independent genes or from introns of protein-coding genes2, 5. In the canonical pathway, primary precursor (pri-miRNA) processing occurs in two steps, catalysed by two members of the RNase III family of enzymes, Drosha and Dicer, operating in complexes with dsRNA-binding proteins (dsRBPs), for example DGCR8 and transactivation-responsive (TAR) RNA-binding protein (TRBP) in mammals. In the first nuclear step, the Drosha–DGCR8 complex processes pri-miRNA into an ~70-nucleotide precursor hairpin (pre-miRNA), which is exported to the cytoplasm. Some pre-miRNAs are produced from very short introns (mirtrons) as a result of splicing and debranching, thereby bypassing the Drosha–DGCR8 step. In either case, cleavage by Dicer, assisted by TRBP, in the cytoplasm yields an ~20-bp miRNA/miRNA* duplex. In mammals, argonaute 2 (AGO2), which has robust RNaseH-like endonuclease activity, can support Dicer processing by cleaving the 3′ arm of some pre-miRNAs, thus forming an additional processing intermediate called AGO2-cleaved precursor miRNA (ac-pre-miRNA)70. Processing of pre-miR-451 also requires cleavage by AGO2, but is independent of Dicer and the 3′ end is generated by exonucleolytic trimming66, 67. Following processing, one strand of the miRNA/miRNA* duplex (the guide strand) is preferentially incorporated into an miRNA-induced silencing complex (miRISC), whereas the other strand (passenger or miRNA*) is released and degraded (not shown). Generally, the retained strand is the one that has the less stably base-paired 5′ end in the miRNA/miRNA* duplex. miRNA* strands are not always by-products of miRNA biogenesis and can also be loaded into miRISC to function as miRNAs26, 58, 59, 60. See Box 2 for details of miRISC function. GW182, glycine-tryptophan protein of 182 kDa; m7G, 7-methylguanosine-cap; PABP, poly(A) binding protein.
, which is exported to the cytoplasm. Some pre-miRNAs are produced from very short introns (mirtrons) as a result of splicing and debranching, thereby bypassing the Drosha–DGCR8 step. In either case, cleavage by Dicer, assisted by TRBP, in the cytoplasm yields an ~20-bp miRNA/miRNA* duplex. In mammals, argonaute 2 (AGO2), which has robust RNaseH-like endonuclease activity, can support Dicer processing by cleaving the 3′ arm of some pre-miRNAs, thus forming an additional processing intermediate called AGO2-cleaved precursor miRNA (ac-pre-miRNA)70. Processing of pre-miR-451 also requires cleavage by AGO2, but is independent of Dicer and the 3′ end is generated by exonucleolytic trimming66, 67. Following processing, one strand of the miRNA/miRNA* duplex (the guide strand) is preferentially incorporated into an miRNA-induced silencing complex (miRISC), whereas the other strand (passenger or miRNA*) is released and degraded (not shown). Generally, the retained strand is the one that has the less stably base-paired 5′ end in the miRNA/miRNA* duplex. miRNA* strands are not always by-products of miRNA biogenesis and can also be loaded into miRISC to function as miRNAs26, 58, 59, 60. See Box 2 for details of miRISC function. GW182, glycine-tryptophan protein of 182 kDa; m7G, 7-methylguanosine-cap; PABP, poly(A) binding protein.")
31
tumour suppressive and oncogenic microRNAs
T Paranjape, et al. GUT. 2009

32
Profiling the mature and primary microRNA transcripts
Drosha Cleavage site Pri-miRNA /~-OH Thomson, J. M/Hammond S

35
Global Reduction in miRNAs in the Context of Cancer…
Red = Abundant Blue= Depleted Lu, J., et al Nature 435:

36
Thomson, et al Genes Dev. 20(16):22202-7 2006
:")
39
Next generation sequencing
miRNA sequencing RNA sequencing HITS-CLIP sequencing

40
miRNA seq short RNAs that are about 21-25bp are first selected by column or electrophoresis. A starting quantity of ug total RNA is required for gel purification and size selection. Adaptor ligation adds DNA adapters to both ends of the small RNAs, which act as primer binding sites during RT and PCR amplification. Then these small adaptor-ligated RNAs will be RT and PCR and then sequencing

41
RNA seq provides information on the level of RNA transcribed from a particular genome, can be use to identify miRNA targetome. Total RNA is first isolated and then Poly A library is constructed by using poly T primer for all the coding RNAs, followed by NGS and transcriptome alignment. Disadvantage: will miss the mRNA targets that are regulated by miRNA at translational repression.

42
HITS-CLIP seq High-throughput sequencing of RNAs isolated by crosslinking immunoprecipitation (HITS-CLIP), is a genome-wide means of mapping protein–RNA binding sites in vivo. UV irradiation to crosslink RNA to associated RNA-binding proteins, then IP using antibody against argonaute protein (AGO2 orAGO1), followed by deep-sequencing. It identifies direct target sequences through the sequencing of RNAs from immuneoprecipitated cross-linked Argonaute-miRNA-mRNA complexes. Starbase is the database for exploring protein-RNA and miRNA-target interactions from HITS-CLIP
, is a genome-wide means of mapping protein–RNA binding sites in vivo. UV irradiation to crosslink RNA to associated RNA-binding proteins, then IP using antibody against argonaute protein (AGO2 orAGO1), followed by deep-sequencing. It identifies direct target sequences through the sequencing of RNAs from immuneoprecipitated cross-linked Argonaute-miRNA-mRNA complexes. Starbase is the database for exploring protein-RNA and miRNA-target interactions from HITS-CLIP.")
43
HITS-CLIP (CLIP-Seq) Thomson D et.al., 2011
 Thomson D et.al., 2011")
44
Non-coding RNAs (ncRNAs)
ncRNAs: A brief intro Long non-coding RNAs (lncRNAs or lincRNAs): >200 bp miRNAs: Biogenesis, measurement, functional analysis, utility Key references: Prensner JR & Chinnaiyan AM. The emergence of lincRNAs in cancer biology. Cancer Disc. 1: , 2011. 2. Guttman M and Rinn JL. Modular regulatory principles of large non-coding RNAs. Nature 482, , 2012. 3. Lee JT. Epigenetic regulation by long noncoding RNAs. Science 338: , 2012. MicroRNAs (miRNAs) are processed from RNA polymerase II (RNAPII)-specific transcripts of independent genes or from introns of protein-coding genes2, 5. In the canonical pathway, primary precursor (pri-miRNA) processing occurs in two steps, catalysed by two members of the RNase III family of enzymes, Drosha and Dicer, operating in complexes with dsRNA-binding proteins (dsRBPs), for example DGCR8 and transactivation-responsive (TAR) RNA-binding protein (TRBP) in mammals. In the first nuclear step, the Drosha–DGCR8 complex processes pri-miRNA into an ~70-nucleotide precursor hairpin (pre-miRNA), which is exported to the cytoplasm. Some pre-miRNAs are produced from very short introns (mirtrons) as a result of splicing and debranching, thereby bypassing the Drosha–DGCR8 step. In either case, cleavage by Dicer, assisted by TRBP, in the cytoplasm yields an ~20-bp miRNA/miRNA* duplex. In mammals, argonaute 2 (AGO2), which has robust RNaseH-like endonuclease activity, can support Dicer processing by cleaving the 3′ arm of some pre-miRNAs, thus forming an additional processing intermediate called AGO2-cleaved precursor miRNA (ac-pre-miRNA)70. Processing of pre-miR-451 also requires cleavage by AGO2, but is independent of Dicer and the 3′ end is generated by exonucleolytic trimming66, 67. Following processing, one strand of the miRNA/miRNA* duplex (the guide strand) is preferentially incorporated into an miRNA-induced silencing complex (miRISC), whereas the other strand (passenger or miRNA*) is released and degraded (not shown). Generally, the retained strand is the one that has the less stably base-paired 5′ end in the miRNA/miRNA* duplex. miRNA* strands are not always by-products of miRNA biogenesis and can also be loaded into miRISC to function as miRNAs26, 58, 59, 60. See Box 2 for details of miRISC function. GW182, glycine-tryptophan protein of 182 kDa; m7G, 7-methylguanosine-cap; PABP, poly(A) binding protein. Dean G. Tang (Molecular Biology of Cancer; 3/6/2013)
: >200 bp. miRNAs: Biogenesis, measurement, functional analysis, utility. Key references: Prensner JR & Chinnaiyan AM. The emergence of lincRNAs in cancer biology. Cancer Disc. 1: , Guttman M and Rinn JL. Modular regulatory principles of large non-coding RNAs. Nature 482, , Lee JT. Epigenetic regulation by long noncoding RNAs. Science 338: , MicroRNAs (miRNAs) are processed from RNA polymerase II (RNAPII)-specific transcripts of independent genes or from introns of protein-coding genes2, 5. In the canonical pathway, primary precursor (pri-miRNA) processing occurs in two steps, catalysed by two members of the RNase III family of enzymes, Drosha and Dicer, operating in complexes with dsRNA-binding proteins (dsRBPs), for example DGCR8 and transactivation-responsive (TAR) RNA-binding protein (TRBP) in mammals. In the first nuclear step, the Drosha–DGCR8 complex processes pri-miRNA into an ~70-nucleotide precursor hairpin (pre-miRNA), which is exported to the cytoplasm. Some pre-miRNAs are produced from very short introns (mirtrons) as a result of splicing and debranching, thereby bypassing the Drosha–DGCR8 step. In either case, cleavage by Dicer, assisted by TRBP, in the cytoplasm yields an ~20-bp miRNA/miRNA* duplex. In mammals, argonaute 2 (AGO2), which has robust RNaseH-like endonuclease activity, can support Dicer processing by cleaving the 3′ arm of some pre-miRNAs, thus forming an additional processing intermediate called AGO2-cleaved precursor miRNA (ac-pre-miRNA)70. Processing of pre-miR-451 also requires cleavage by AGO2, but is independent of Dicer and the 3′ end is generated by exonucleolytic trimming66, 67. Following processing, one strand of the miRNA/miRNA* duplex (the guide strand) is preferentially incorporated into an miRNA-induced silencing complex (miRISC), whereas the other strand (passenger or miRNA*) is released and degraded (not shown). Generally, the retained strand is the one that has the less stably base-paired 5′ end in the miRNA/miRNA* duplex. miRNA* strands are not always by-products of miRNA biogenesis and can also be loaded into miRISC to function as miRNAs26, 58, 59, 60. See Box 2 for details of miRISC function. GW182, glycine-tryptophan protein of 182 kDa; m7G, 7-methylguanosine-cap; PABP, poly(A) binding protein. Dean G. Tang (Molecular Biology of Cancer; 3/6/2013)")
55
How do miRNAs regulate tumorigenic PCa cells?
Can (Julie) Liu PCa stem/progenitor cells (AR-/lo; PSA-/lo) Holoclones ALDHhi CD44+ a2b1+ SP (PSA+/hi) ABCG2+ Our experiments with xenografts and cell lines have given us tremendous insight into the id of tumorigenic cells in pca, but the truth is that they are different from primary human prostate tumors and in order to apply our findings to a clinical setting, we need to first see if our findings can be reproduced using primary human adenocarcinomas. Increasingly ‘mature’ PCa cells Prolif.
 Liu. PCa stem/progenitor cells. (AR-/lo; PSA-/lo) Holoclones. ALDHhi. CD44+ a2b1+ SP. (PSA+/hi) ABCG2+ Our experiments with xenografts and cell lines have given us tremendous insight into the id of tumorigenic cells in pca, but the truth is that they are different from primary human prostate tumors and in order to apply our findings to a clinical setting, we need to first see if our findings can be reproduced using primary human adenocarcinomas. Increasingly ‘mature’ PCa cells. Prolif.")
56
miR-34a is Underexpressed in CD44+ PCa Cells
56

57
Ectopic Expression of miR-34a Inhibits Prostate Tumor Development
57

58
Re-expression of miR-34a in CD44+ PCa cells abolishes tumor regeneration
58

59
Anti-miR-34a Promotes PCa Regeneration and Metastasis
59

60
Systemically delivered miR-34a inhibits tumor development and metastasis
PC3 Therapeutic Exp NC miR-34a Not difference in metastasis, explaination? 60

61
CD44 as a DIRECT & FUNCTIONAL target of miR-34a
Note tumor weights not that much different because the tumors are not harvested at the same time. 61

62
CD44 knockdown phenocopies miR-34a effects
Our experiments with xenografts and cell lines have given us tremendous insight into the id of tumorigenic cells in pca, but the truth is that they are different from primary human prostate tumors and in order to apply our findings to a clinical setting, we need to first see if our findings can be reproduced using primary human adenocarcinomas.

63
CD44 knockdown phenocopies miR-34a effects
Our experiments with xenografts and cell lines have given us tremendous insight into the id of tumorigenic cells in pca, but the truth is that they are different from primary human prostate tumors and in order to apply our findings to a clinical setting, we need to first see if our findings can be reproduced using primary human adenocarcinomas.

64
Our experiments with xenografts and cell lines have given us tremendous insight into the id of tumorigenic cells in pca, but the truth is that they are different from primary human prostate tumors and in order to apply our findings to a clinical setting, we need to first see if our findings can be reproduced using primary human adenocarcinomas.

65
Overexpression of miR-17-19b in a Mouse Model of Human Lymphoma
Thomson, J. M/Hammond S

66
Expression of miR17-19b Results in B-cell Lymphoma
He, et al Nature 435(7043):
:")
67
miRNA as therapeutics company Microrna disease formula Half life
Regulus Anti-21, anti-122, Anti-10b HCC Hepatitis C GBM Single stranded modified oligos 21 days stability does not leads to destruction of miRNA. Santaris Pharma Miravirsen (anti-21) LNA 5 doses in 120 days, 2weeks stability Phase ii, first in human miRgen Anti-miR-208 Heart failure Alternative week injection Reverse heart failure (induced by high Na+ diet) miRx miR-34a Liver cancer (Hep3B cell injection) Oligo+Nov340 Pre-clinical Target HDAC1
 LNA. 5 doses in 120 days, 2weeks stability. Phase ii, first in human. miRgen. Anti-miR-208. Heart failure. Alternative week injection. Reverse heart failure (induced by high Na+ diet) miRx. miR-34a. Liver cancer (Hep3B cell injection) Oligo+Nov340. Pre-clinical. Target HDAC1.")
68
MicroRNAs (miRNAs) are processed from RNA polymerase II (RNAPII)-specific transcripts of independent genes or from introns of protein-coding genes2, 5. In the canonical pathway, primary precursor (pri-miRNA) processing occurs in two steps, catalysed by two members of the RNase III family of enzymes, Drosha and Dicer, operating in complexes with dsRNA-binding proteins (dsRBPs), for example DGCR8 and transactivation-responsive (TAR) RNA-binding protein (TRBP) in mammals. In the first nuclear step, the Drosha–DGCR8 complex processes pri-miRNA into an ~70-nucleotide precursor hairpin (pre-miRNA), which is exported to the cytoplasm. Some pre-miRNAs are produced from very short introns (mirtrons) as a result of splicing and debranching, thereby bypassing the Drosha–DGCR8 step. In either case, cleavage by Dicer, assisted by TRBP, in the cytoplasm yields an ~20-bp miRNA/miRNA* duplex. In mammals, argonaute 2 (AGO2), which has robust RNaseH-like endonuclease activity, can support Dicer processing by cleaving the 3′ arm of some pre-miRNAs, thus forming an additional processing intermediate called AGO2-cleaved precursor miRNA (ac-pre-miRNA)70. Processing of pre-miR-451 also requires cleavage by AGO2, but is independent of Dicer and the 3′ end is generated by exonucleolytic trimming66, 67. Following processing, one strand of the miRNA/miRNA* duplex (the guide strand) is preferentially incorporated into an miRNA-induced silencing complex (miRISC), whereas the other strand (passenger or miRNA*) is released and degraded (not shown). Generally, the retained strand is the one that has the less stably base-paired 5′ end in the miRNA/miRNA* duplex. miRNA* strands are not always by-products of miRNA biogenesis and can also be loaded into miRISC to function as miRNAs26, 58, 59, 60. See Box 2 for details of miRISC function. GW182, glycine-tryptophan protein of 182 kDa; m7G, 7-methylguanosine-cap; PABP, poly(A) binding protein.
, which is exported to the cytoplasm. Some pre-miRNAs are produced from very short introns (mirtrons) as a result of splicing and debranching, thereby bypassing the Drosha–DGCR8 step. In either case, cleavage by Dicer, assisted by TRBP, in the cytoplasm yields an ~20-bp miRNA/miRNA* duplex. In mammals, argonaute 2 (AGO2), which has robust RNaseH-like endonuclease activity, can support Dicer processing by cleaving the 3′ arm of some pre-miRNAs, thus forming an additional processing intermediate called AGO2-cleaved precursor miRNA (ac-pre-miRNA)70. Processing of pre-miR-451 also requires cleavage by AGO2, but is independent of Dicer and the 3′ end is generated by exonucleolytic trimming66, 67. Following processing, one strand of the miRNA/miRNA* duplex (the guide strand) is preferentially incorporated into an miRNA-induced silencing complex (miRISC), whereas the other strand (passenger or miRNA*) is released and degraded (not shown). Generally, the retained strand is the one that has the less stably base-paired 5′ end in the miRNA/miRNA* duplex. miRNA* strands are not always by-products of miRNA biogenesis and can also be loaded into miRISC to function as miRNAs26, 58, 59, 60. See Box 2 for details of miRISC function. GW182, glycine-tryptophan protein of 182 kDa; m7G, 7-methylguanosine-cap; PABP, poly(A) binding protein.")
Similar presentations















>")

 microRNAs 11/6 2009 Peter Hagedorn.>")

 are a class of small RNA molecules, about ~21 nucleotide (nt) long. MicroRNA are small non coding RNAs (ncRNAs) that regulate.>")

>")

>")




