Download presentation
Presentation is loading. Please wait.

1
Chapter 9 DNA REPLICATION
The replication or duplication of DNA depends on one main idea, namely that the nitrogen bases of the nucleotides are complementary to each other on opposite halves of the molecule . Thus the base adenine(A) in DNA always pairs with thymine(T) and guanine(G) with cytosine(C).
 in DNA always pairs with thymine(T) and guanine(G) with cytosine(C).")
2
This pairing is due to the shapes of the nucleotide bases which allow hydrogen bonds to form between the complementary bases, holding the two halves of the DNA molecule together. The parent DNA molecule unzips exposing the two halves of the DNA molecule .

3
DNA REPLICATION Each half of the parent DNA molecule serves as a template for the complementarity bases to be brought into the correct position to make a new complementary half for each of the original parent halves. This theoretical consideration suggested to Watson andCrick that DNA replication is what we now call semiconservative

4
The Watson-Crick model included three possibilities of DNA replication
Semiconservative (proven to be true) Conservative Dispersive Semi conservative DNA replication means that when the 2 new DNA molecules are formed during DNA replication, each resulting daughter molecule consists of one side from the original parent molecule and a new side synthesized from the parent side which served as a template.
 Conservative. Dispersive. Semi conservative DNA replication means that when the 2 new DNA molecules are formed during DNA replication, each resulting daughter molecule consists of one side from the original parent molecule and a new side synthesized from the parent side which served as a template.")
5
Experiment of Matthew Meselson and Frank Stahl demonstrated that DNA replication was semi-conservative. These scientists grew the bacteria E-coli in media containing nutrients with the isotope Nitrogen 15, 15N as opposed to the normal isotope of nitrogen, 14N. After a time some of the DNA containing the normal isotope is replaced with DNA containing ,15N as new DNA synthesis proceeds.

6
They cultured the bacteria for a sufficient amount of time that so that they could be sure that all the DNA contained just 15N. Then they took these bacteria and set up cultures using growth medium containing 14N and followed the bacteria through a number of replication cycles. Meselson and Stahl then knew they could separate DNA's with differing amounts of 15N by using a process called cesium density gradient centrifugation which separates molecules by weight in a solution of cesium chloride. These scientists noted that one would expect a different distribution of DNA's by weight depending on whether the type of replication involved.

7
For example DNA replication could be conservative, in which case after one replication cycle half the DNA would contain only 14N and thus be lighter and the other half of the DNA would contain only 15N and thus be heavier. Thus there would be two DNA bands, one light and one heavy produced by the centrifugation.

8
DNA replication could have been dispersive in which case the daughter molecules after one generation would contain roughly equal mixtures of 15N and 14N mixed through each of the halves of the molecule, and there would only be a single DNA band of intermediate weight.

9
Before a cell can divide, it must duplicate all its DNA.
In eukaryotes, this occurs during S phase of the cell cycle. A portion of the double helix is unwound by a helicase. DNA Helicase attaches at initiation site. DNA Helicase is an enzyme that unravels the DNA double helix and breaks the hydrogen bonds. A separate set of proteins prevents the two halves of the DNA molecule from forming hydrogen bonds or reannealing where replication is beginning.

10
A molecule of a DNA polymerase binds to one strand of the DNA and begins moving along it in the 3' to 5' direction, using it as a template for assembling a leading strand of nucleotides and reforming a double helix. In eukaryotes, this molecule is called DNA polymerase delta (δ).
.")
11
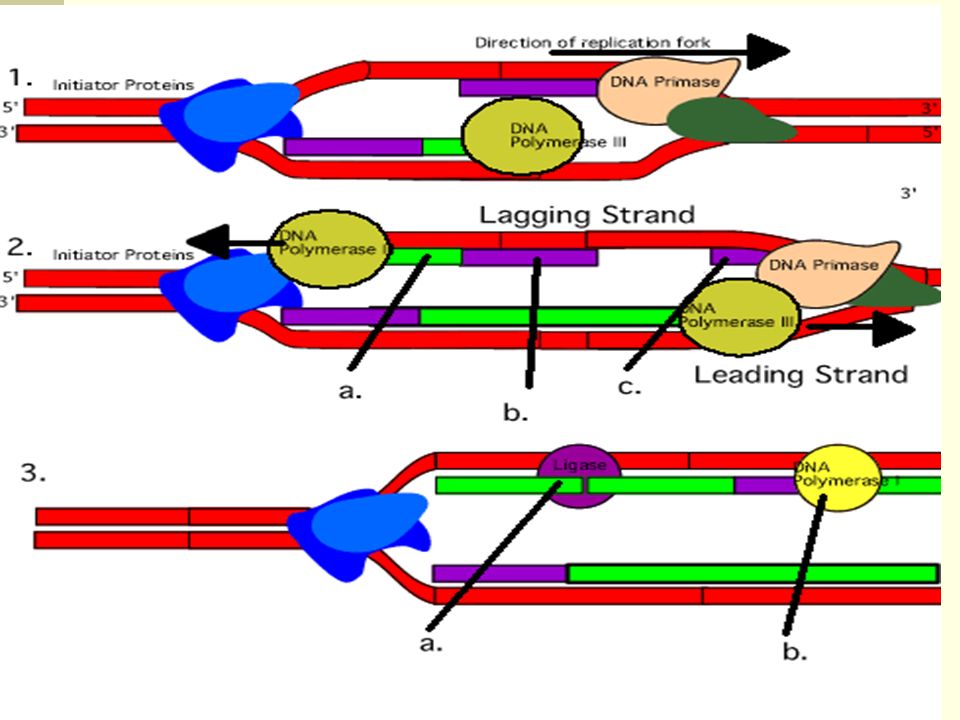
Because DNA synthesis can only occur 5' to 3', a molecule of a second type of DNA polymerase (epsilon, ε, in eukaryotes) binds to the other template strand as the double helix opens. This molecule synthesize discontinuous segments of polynucleotides (called Okazaki fragments). Another enzyme, DNA ligase then stitches these together into the lagging strand.
. Another enzyme, DNA ligase then stitches these together into the lagging strand.")
12
Step 1: 30 copies of initiator protein bind
to a specic DNA site (OriC)
")
14
Initiation and termination of DNA replication
Step 1: 30 copies of initiator protein bind to a specic DNA site (OriC) Step 2: DNA wraps around the initiator protein complex, which induces local separation of DNA strands at AT-rich DNA site (formation of two replication forks) Step 3: Single strand binding proteins (SSB) bind to single DNA strands, and primasomes enter the forks (one per fork) Step 4: Other components of DNA replication are assembled and replication begins Step 5: The two replication forks meet at the opposite side of the DNA, in the region termed ter Step 6: The replication enzymes disengage, the two DNA (double stranded) molecules separate. The later process is mediated by a special topoisomerase
 Step 2: DNA wraps around the initiator protein complex, which induces local separation of DNA strands at AT-rich DNA site (formation of two replication forks) Step 3: Single strand binding proteins (SSB) bind to single DNA strands, and primasomes enter the forks (one per fork) Step 4: Other components of DNA replication are assembled and replication begins. Step 5: The two replication forks meet at the opposite side of the DNA, in the region termed ter. Step 6: The replication enzymes disengage, the two DNA (double stranded) molecules separate. The later process is mediated by a special topoisomerase.")
15
The structure and functions of DNA polymerases
Polymerase I – DNA repair; removes RNA primer in Okazaki fragments and replaces them with DNA Polymerase II – unclear function Polymerase III – Synthesizes DNA strands Polymerase III is a part of large complex – DNA polymerase III holoenzyme; one of the subunits of this holoenzyme is ß-subunit, which keeps the polymerase III associated with DNA

16
Exonuclease activities of DNA polymerase
All DNA polymerases possess exonuclease activity Exonuclease – an enzyme that degrades nucleic acids by removing one terminal nucleotide at a time There are 5’à 3’ and 3’à 5’ exonucleases DNA polymerase I can act as 5’à 3’ and 3’à 5’ exonuclease. The role of 5 à 3’ exonuclease of DNA polymerase I Can degrade both DNA and RNA Removes the RNA primer in the Okazaki fragment; simultaneously, the polymerase activity of this enzyme fills the gap with DNA nucleotides The role of 3’à 5’ exonuclease of DNA polymerase All three DNA polymerses possess 3’à 5’ exonuclease activity Removes mismatched nucleotides from DNA strands during DNA replication – immediate proofreading, and replaces "wrong" nucleotides with the "correct" ones (polymerase III) Postreplicative mismatch repair – corrects mismatched nucleotides after DNA replication
 Postreplicative mismatch repair – corrects mismatched nucleotides after DNA replication.")
17
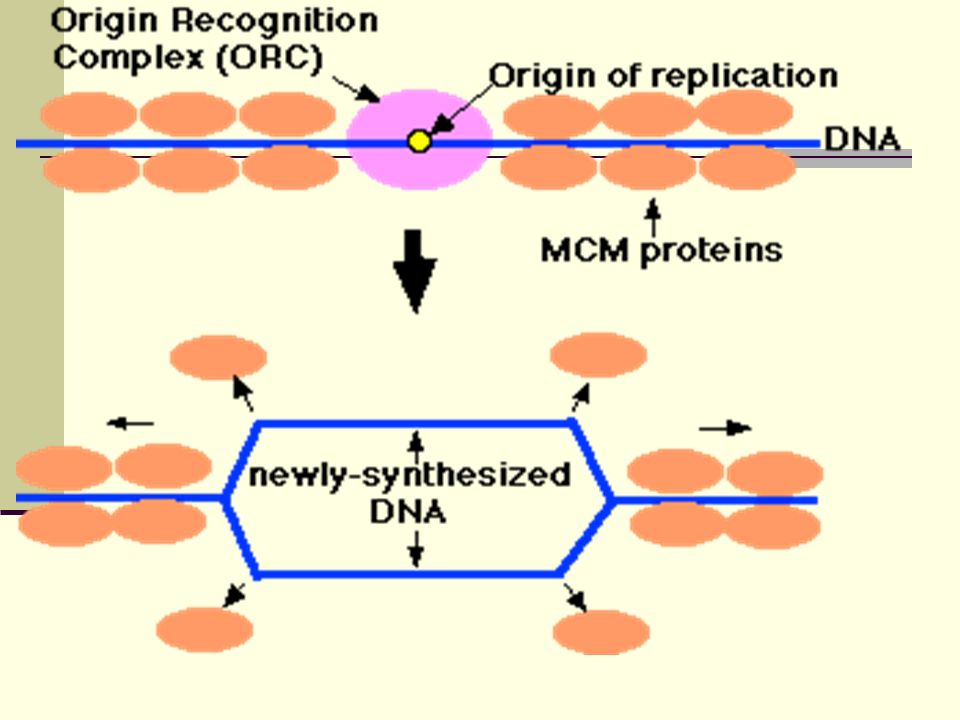
Replicons, autonomous replication sequences,
Replicons located close together tend to replicate simultaneously Replication is initiated at the DNA site that contains specific sequences – autonomous replication sequences (ARS) ARS are scattered throughout DNA A protein complex – origin recognition complex (ORC) – binds to ARS; remains bound throughout the cell cycle Replication is triggered by the binding of other proteins to ORC
 ARS are scattered throughout DNA. A protein complex – origin recognition complex (ORC) – binds to ARS; remains bound throughout the cell cycle. Replication is triggered by the binding of other proteins to ORC.")
18
Restricting replication to once per cell cycle
Step 1: The origin of replication (ARS) is bound to origin recognition complex (ORC) Step 2: Proteins, licesing factors, bind to ARS and switch them into a state that is competent for replication; licesing factors can only gain access to the chromosomes during mitosis when the nuclear envelope does not exist Step 3: An activation factor binds to the chromosome and induces licensed origins to initiate replication
 is bound to origin recognition complex (ORC) Step 2: Proteins, licesing factors, bind to ARS and switch them into a state that is competent for replication; licesing factors can only gain access to the chromosomes during mitosis when the nuclear envelope does not exist. Step 3: An activation factor binds to the chromosome and induces licensed origins to initiate replication.")
19
Speed of Replication Prokaryotes
The single molecule of DNA that is the E. coli genome contains 4.7 x 106 nucleotide pairs. DNA replication begins at a single, fixed location in this molecule, the replication origin, proceeds at about 1000 nucleotides per second, and thus is done in no more than 40 minutes. E. coli genome (4.7 x 106) is copied without error!
 is copied without error!")
20
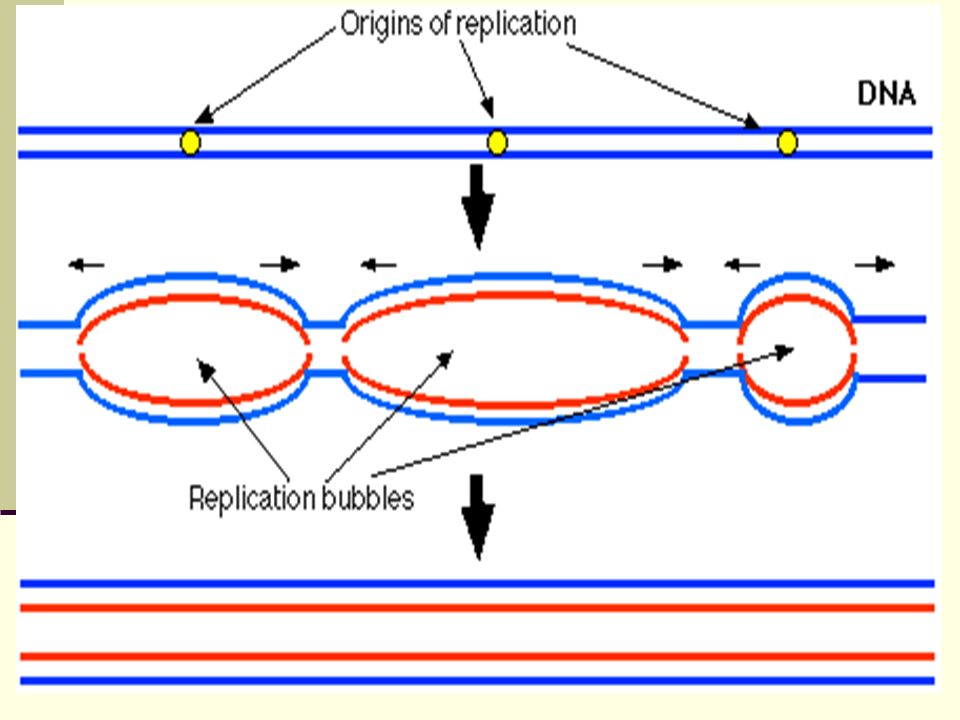
Eukaryotes The average human chromosome contains 150 x 106 nucleotide pairs which are copied at about 50 base pairs per second. The process would take a month (rather than the hour it actually does) but for the fact that there are many places on the eukaryotic chromosome where replication can begin. Replication begins at some replication origins earlier in S phase than at others, but the process is completed for all by the end of S phase. As replication nears completion, "bubbles" of newly replicated DNA meet and fuse, finally forming two new molecules.
 but for the fact that there are many places on the eukaryotic chromosome where replication can begin. Replication begins at some replication origins earlier in S phase than at others, but the process is completed for all by the end of S phase. As replication nears completion, bubbles of newly replicated DNA meet and fuse, finally forming two new molecules.")
22
Bidirectional DNA Replication in Eukaryotic Cells
DNA replication (arrows) occurs in both directions from multiple origins of replication in the linear DNA found in eukaryotic cells.
 occurs in both directions from multiple origins of replication in the linear DNA found in eukaryotic cells.")
25
Solving the problem of overwound DNA
Enzymes, topoisomerases, can change the state of supercoiling in a DNA molecule One topoisomerase, DNA gyrase,travels along the DNA ahead of the replication fork The enzyme changes the positively supercoiled DNA into negatively supercoiled DNA by cleaving the strands and then sealing the cuts

26
DNA polymerases Enzymes that synthesize new DNA strands
DNA polymerase I DNA polymerase II DNA polymerase III DNA polymerase moves along a template DNA strand and incorporates a complementary nucleotide onto the end of the strand being assembled Templates (b) and nontemplates (a) for DNA polymerase activity DNA polymerase can only add nucleotides to the 3’ hydroxil (OH) terminus of an existing strand The strand that provides 3’OH terminus is called a primer
 and nontemplates (a) for DNA polymerase activity. DNA polymerase can only add nucleotides to the 3’ hydroxil (OH) terminus of an existing strand. The strand that provides 3’OH terminus is called a primer.")
27
Primasome and DNA polymerase III
Primasome – a complex composed of a primase and DNA helicase Primasome moves along the lagging-strand template As the helicase unwinds the DNA (opens DNA strands), primase synthesizes RNA primers RNA primers are subsequently extended by a DNA polymerase III (synthesis of Okazaki fragments) The same DNA polymerase III synthesizes successive Okazaki fragmentsReplisome
, primase synthesizes RNA primers. RNA primers are subsequently extended by a DNA polymerase III (synthesis of Okazaki fragments) The same DNA polymerase III synthesizes successive Okazaki fragmentsReplisome.")
28
Replisome The DNA polymerase III that is moving along the lagging template is bound to the DNA polymerase III that is moving along the leading template – complex called replisome Lagging DNA strand forms a loop so that two polymerases can move together without violating "5’à 3’ rule" for DNA replication

Similar presentations



 Two strands of DNA run antiparallel.>")

 and Replication of DNA (chapter 12, pages 318 – 334)>")





 – Examples: Humans begin as a single fertilized.>")







. This structure has novel.>")
