Download presentation
Presentation is loading. Please wait.

1
Lecture 7 Brief review of enzyme mechanisms and kinetics
A ‘toolbox’ for mechanistic biochemists. A path to the molecule: protein purification techniques.

2
Protein Kinase A spatially organizes ATP and peptide chain to facilitate the reaction

3
kB – Boltzmann constant
Rate of ‘hopping’ over the barrier: kB – Boltzmann constant h – Plank constant frequency pre-factor Boltzmann factor DG*s→p DG*p→s DGo

4
Binding of a molecule to the catalyst reduces the energy of transition state
AA = A+A AA Binding energy A+A In the bound state to enzyme I the bond is relaxed (no catalysis) In the bound state to enzyme II the bond is stretched
 In the bound state to enzyme II the bond is stretched.")
5
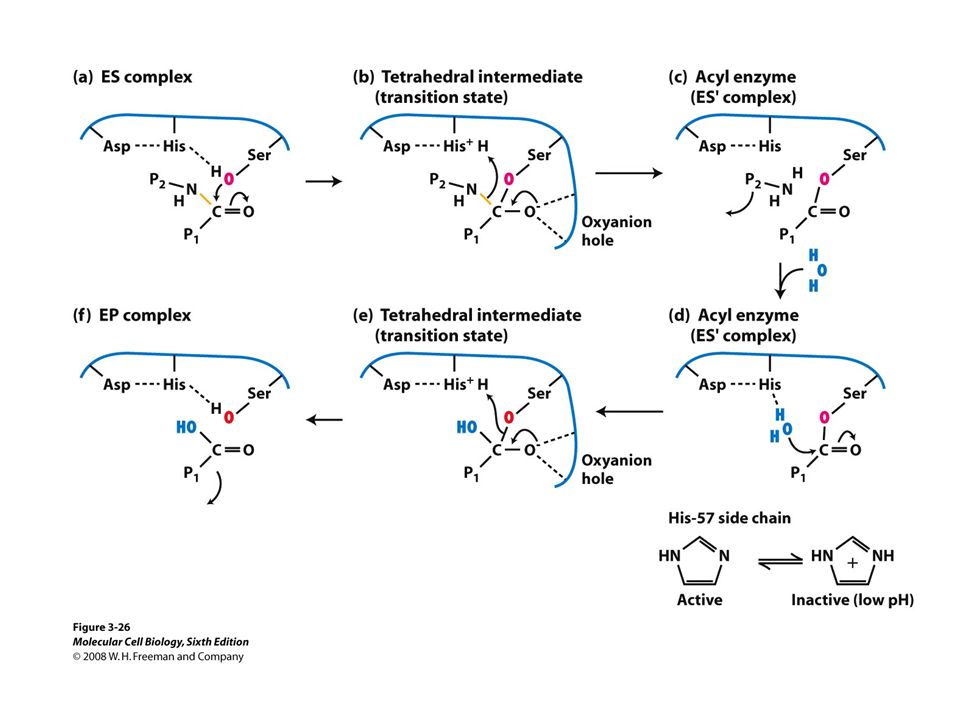
Trypsin: the binding pocket and the catalytic site

6
His58 Asp102 Ser195 Asp189

7
His58 Asp102 Ser195 Asp189

8
His-58

9
The sidechain specificity pocket
cuts after Arg (R), Lys (K) cuts after Tyr (Y), Trp (W) or Phe (F) cuts after any small side-chain residue Gly (G), Ala (A), Ser (S)
, Lys (K) cuts after. Tyr (Y), Trp (W) or Phe (F) cuts after. any small side-chain residue Gly (G), Ala (A), Ser (S)")
11
Catalysis involves simple binding
Langmuir

12
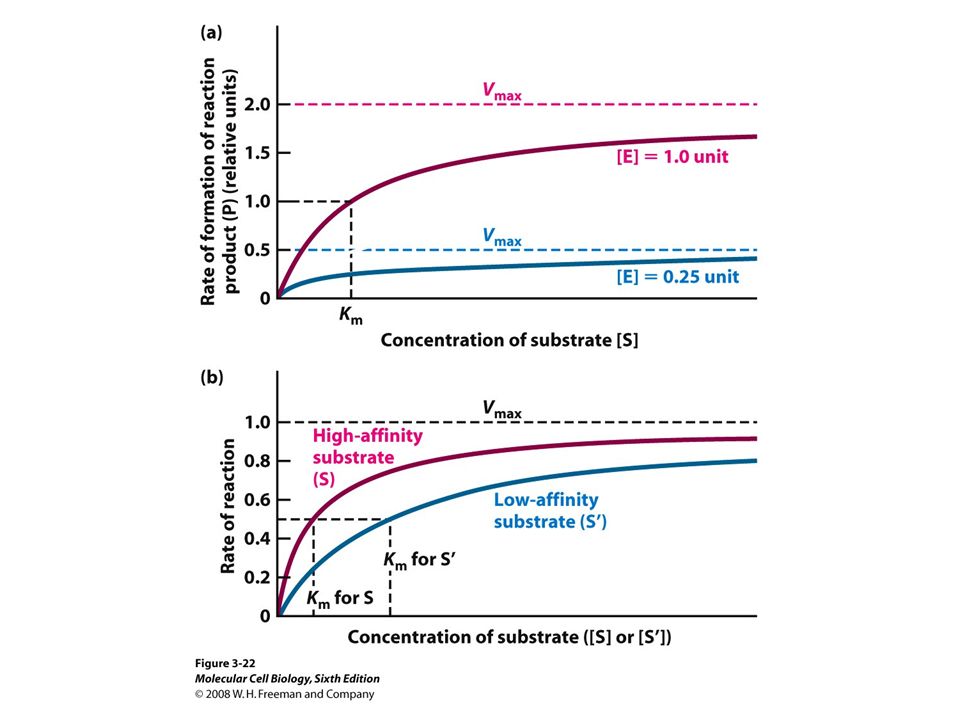
Michaelis-Menten equation for enzyme/transport reactions is very similar to the Langmuir isotherm
[s], mM 10 20 30 40 50 0.2 0.4 0.6 0.8 1 Km = 1 mM Km = 10 mM Vmax A “simple explanation” says that the rate of reaction should be proportional to the occupancy of the binding site as long as Vmax is constant.

13
E + S ↔ ES ↔ EP ↔ E + P DG*uncat E+S DG*cat E+P
Rate of ‘hopping’ over the barrier: DG*uncat E+S DG*cat E+P E + S ↔ ES ↔ EP ↔ E + P

14
E + S ↔ ES → E + P Et – total enzyme k1 k2 k-1 At steady state:
(we assume k-2 = 0 ) k-1 Et – total enzyme Rate of ES formation Rate of ES breakdown At steady state: Michaelis-Menten again
 k-1. Et – total enzyme. Rate of ES formation. Rate of ES breakdown. At steady state: Michaelis-Menten again.")
16
How do we know who are the players in the structural assembly, reaction of interest or specific signaling cascade? 1. Standard biochemical ‘brute-force’ approach: isolate/purify the protein component(s) and show that they are functional or necessary for the specific function; then clone the gene(s), mutate or knock them out (down) and demonstrate altered function (biochemistry+reverse genetics). 2. Genetic approach: mutagenize and see the phenotypic deviation, find the mutant, identify and clone the gene conferring that specific trait, predict its product, generate the knock-out and show the interdependence between the phenotype and the presence of intact gene coding for that protein (forward genetics). 3. A reductionistic finale of either path toward the molecule: for most of the mechanistic studies it is important to express and isolate that protein, find the conditions under which it is active in isolation, reconstitute its functional state in a test tube and determine its properties in vitro. Obtain structural and mechanistic clues on how the molecular machine works.
 and show that they are functional or necessary for the specific function; then clone the gene(s), mutate or knock them out (down) and demonstrate altered function (biochemistry+reverse genetics). 2. Genetic approach: mutagenize and see the phenotypic deviation, find the mutant, identify and clone the gene conferring that specific trait, predict its product, generate the knock-out and show the interdependence between the phenotype and the presence of intact gene coding for that protein (forward genetics). 3. A reductionistic finale of either path toward the molecule: for most of the mechanistic studies it is important to express and isolate that protein, find the conditions under which it is active in isolation, reconstitute its functional state in a test tube and determine its properties in vitro. Obtain structural and mechanistic clues on how the molecular machine works.")
17
Cell disruption and fractionation

19
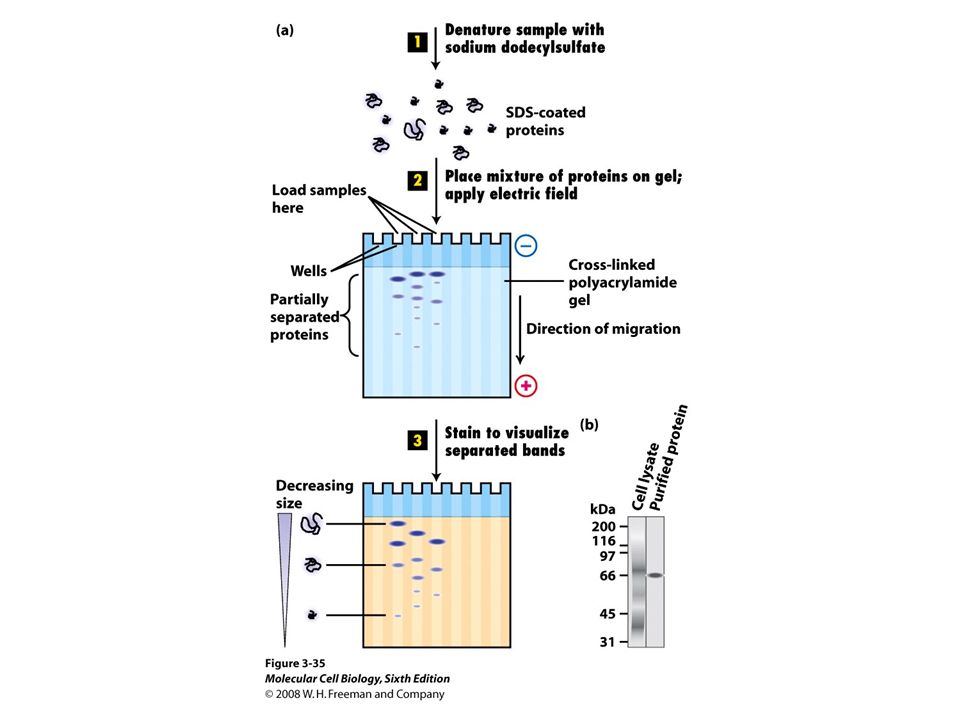
2D gels separate first by charge an then by size

20
Small molecules have longer diffusion distance because they penetrate the beads

23
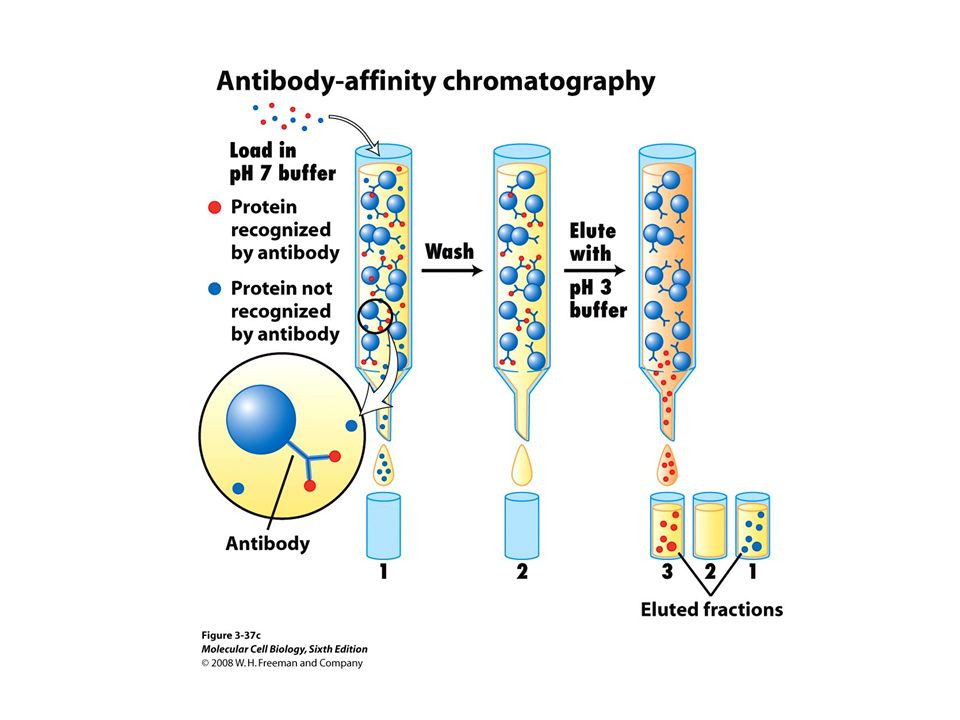
Western Blotting

24
Mass spectrometry measures m/z (mass/charge)
")
25
Complete sequencing of a protein can be accomplished using MS/MS in conjunction with genomic information

Similar presentations




![Review: Amino Acid Side Chains Aliphatic- Ala, Val, Leu, Ile, Gly Polar- Ser, Thr, Cys, Met, [Tyr, Trp] Acidic (and conjugate amide)- Asp, Asn, Glu, Gln.](/15/4531817/big_thumb.jpg "Review: Amino Acid Side Chains Aliphatic- Ala, Val, Leu, Ile, Gly Polar- Ser, Thr, Cys, Met, [Tyr, Trp] Acidic (and conjugate amide)- Asp, Asn, Glu, Gln.>")



 between.>")

 Involvement.>")


 Examples of Ligand-protein interactions Antibodies and their.>")



 zImpact of genome projects (human, bacteria,>")


