Download presentation
Presentation is loading. Please wait.

1
Recurring conformation of the human immunodeficiency virus type 1 gp120 V3 loop. Stanfield RL, Ghiara JB, Ollmann Saphire E, Profy AT, and Wilson IA. Virology (2003) Oct 10; 315(1) BioInformatics Lab Tuesday, March 9, 2010 Kristoffer Chin Salomon Garcia Michael Piña

2
Outline Introduction Materials and Methods Results and Discussion
Background on HIV Structure Background on HIV V3 Region Materials and Methods Crystallography Results and Discussion References

3
HIV has an outer surface of Glycoproteins
gp120 responsible for binding to CD4 receptors and coreceptors, CCR5 or CXCR4 gp41 anchors gp120 to the viral membrane providing noncovalent association

4
Glycoproteins binding to CD4 T-cells
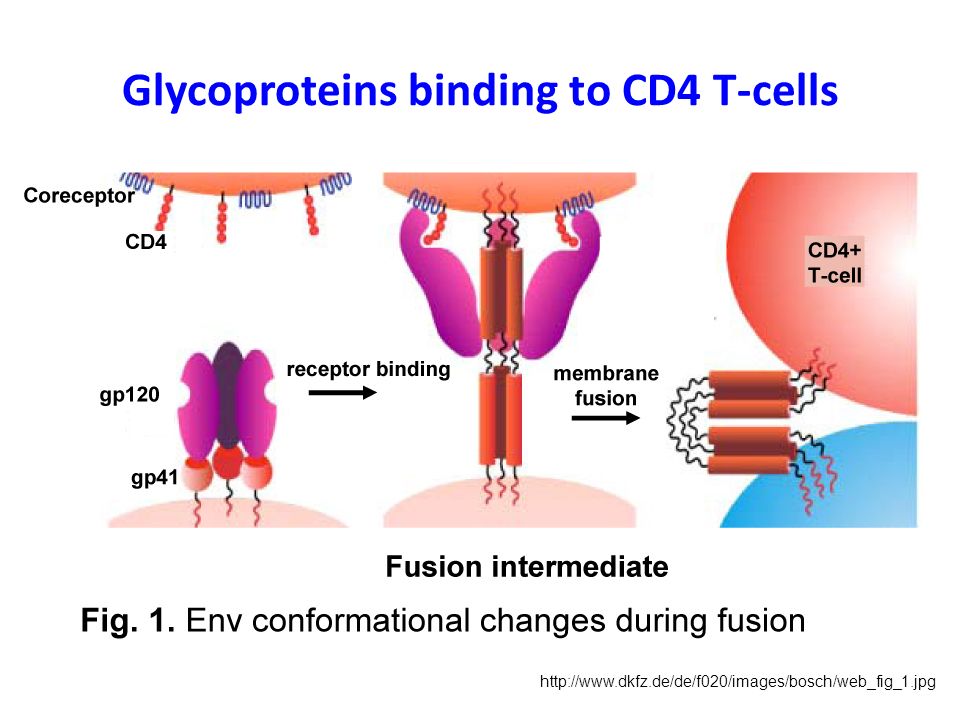
5
V3 Region as part of gp120 V3 important to viral infectivity
V3 important to viral infectivity Progresses initial infection to AIDS

6
Introduction to the Stanfield et al. (2003) Study
V3 regions have high propensity to elicit neutralizing antibodies V3 region inaccessible due to carbohydrates masking or tertiary or quaternary interactions with gp120 complex Is there a limited range of conformational states that gp120 can adopt? How is V3 loop recognized by antibodies and how an alteration of sequence, conformation, or exposure can affect it

7
Finding a dominant conformation for V3
Fabs are antibodies that bind to V3 conformation Fab 83.1, 50.1, and 59.1 bind to a similar conformation of V3 region NMR studies have shown V3 to form similar hairpin loops Stabilization of V3 loops to prevent change of conformation through turns 5 antibodies used for neutralizing and stablization

8
Materials and Methods Mab 83.1 was made by immunization of an ASW mice with cyclic peptide RP70 Antibody was produced in ascites fluid of a mice and purified with an immobilized protein A column Fab was made from immunoglobin by cleavage Fab was concentrated to 15.0 mg/ml for crystallization studies

9
Fab Purification Fab was mixed with 16-mer peptide MP1 in a 6:1 mole ration Crystals were grown using sitting-drop vapor diffusion method with a reservoir solution of 1.6 M Na/K phosphate, 5% isopropanol, pH 6.0. Crystals grow as clusters of thin plates Crystals used for this experiments grew over a 2-week period

10
Fab Crystallization procedure
Crystals were cryocooled to liquid nitrogen temperatures in order to collect the data in a rapid manner The crystals were protected by putting them in a solution containing the following 25% glycerol, 1.6 M Na/K phosphate, 5% isopropanol, pH 6.0 HKL2000 was used to format all of the data that was obtained

11
Structure determination
Matthew coefficient was obtained by two Fab molecules Model was constructed from the constant region of Fab 58.2 EPMR program was used to position the model in the cell EPMR also used to locate the first Fab molecule in the asymetric unit

12
Stanfield et al

13
Model building and refinement
TOM/FRODO was used to rebuild the mutated hybrid model and to correct the sequence and were subsequently refined with CNS version 1.1 Refinement was carried with tight NCS restraints in the beginning and progressively released towards the end of refinement

14
Structural Analysis Kabat convention was used to number the molecules
Light and heavy chains are labeled using “L” and “H” Peptide labeled “P” and was numbered according to HXB2 isolate sequence HBPLUS was used to evaluate the Hydrogen bonds Contacsym program was used to assign van der waals contacts

15
Results and Discussion
Rcryst and Rfree values were slightly higher than other structures determined at 2.6Å resolution Electron density maps were good quality However, repeated refinement and manual rebuilding of the structures caused the higher R-values Rcryst 28.8%, Rfree 32.6% An index (0.45*l) close to an integer value is strong, whereas close to one-half integer is weak
 close to an integer value is strong, whereas close to one-half integer is weak.")
16
Stanfield et al

17
Figure 1: comparison of V3 peptide conformations determined by X-Ray crystallography and NMR
Stanfield et al

18
Figure 2: stereoview of electron density for the V3 peptide bound to Fab 83.1
Stanfield et al

19
The L1 CDR loops have a 5 amino acid insertion after residue L27
All CDR loops fall into their expected canonical classes with the exception of L1 The L1 CDR loops have a 5 amino acid insertion after residue L27 In both Fabs, the tip of this loop bends away from the antigen binding site in an unusual manner Comparison with other L1 loops shows the angle is about 9Å

20
Figure 3: the structure of the Fab 83.1-V3 peptide complex
Stanfield et al

21
Figure 4: Noncanonical loops from Fab 83.1
Stanfield et al

22
CDR H3 has a “kinked” base
This was not predicted from its sequence At least two other Fabs have kinked H3 bases that were not predicted AspH101 normally forms a salt bridge (with Arg or Lys), but in this case it does not which is unexpected
, but in this case it does not which is unexpected.")
23
Stanfield et al

24
Figure 5: stereoview of H3 loop conformations from Fabs 83
Figure 5: stereoview of H3 loop conformations from Fabs 83.1, B02C11, and 26-10 Stanfield et al

25
The peptide makes contact with both the light and heavy chains from the Fab
110 total contacts for 1 molecule 7 are hydrogen bonds with no charge-charge interactions 6 hydrogen bonds are to peptide main-chain atoms 1 bond to Arg side chain The H3 CDR makes the most contacts

26
Stanfield et al

27
Analysis of the 4 peptide reveals that 3 are very similar
The 83.1 peptide structure is the 4th crystal structure determined for a neutralizing antibody V3 peptide complex Analysis of the 4 peptide reveals that 3 are very similar The 4th differs around the V3 region The 4 antibody peptides were generated from related mice The antibodies themselves do not have structural homology The similarity among the conformation of the peptides is not due to the similarity of the Fabs

28
Figure 6: comparison of the V3 peptide conformations
Stanfield et al

29
Figure 7: the antibody combining site of Fab 83.1
Stanfield et al

30
Figure 8: antigen binding sites of Fabs 83.1, 59.1, 58.2, & 50.1
Stanfield et al

31
The peptides, although adopting the same shapes, bind in different orientations and locations in the antibody The antibodies were chosen for ability to neutralize (bind to intact viruses) These peptide conformations should reflect “preferred” conformations of the V3 loop The identified V3 structures represent a recurring conformer on the intact virus
 These peptide conformations should reflect preferred conformations of the V3 loop. The identified V3 structures represent a recurring conformer on the intact virus.")
32
The X-rays of V3 peptides in complex with antibodies help define the range of V3 conformation
Studies suggest that V3 interacts with coreceptors CCR5 and CXCR4 during cell entry This information may be useful in the design of V3 based inhibitors Ultimately, a better understanding of the gp120/gp41 structure (and the V3 region) is vital for understanding how HIV-1 carries out its binding and fusion activities
 is vital for understanding how HIV-1 carries out its binding and fusion activities.")
33
References Stanfield et al. Recurring conformation of the human immunodeficiency virus type 1 gp120 V3 loop. Virology (2003) Oct 10; 315(1)

Similar presentations
 PROTEIN.>")










 A functional proteins consists of one or more polypeptides that have been precisely twisted,>")







