Download presentation
Presentation is loading. Please wait.

1
Welcome to Our Microbial Genetics Class
Lesson Four College of Bioengineering Tianjin University of Science and Technology

2
C H A P T E R 13 Microbial Recombination and Plasmids
Concepts Recombination is a one-way process in procaryotes: a piece of genetic material (the exogenote) is donated to the chromosome of a recipient cell (the endogenote) and integrated into it. 2. The actual transfer of genetic material between bacteria usually takes place in one of three ways: direct transfer between two bacteria temporarily in physical contact (conjugation), transfer of a naked DNA fragment (transformation), or transport of bacterial DNA by bacteriophages (transduction). 3. Plasmids and transposable elements can move genetic material between bacterial chromosomes and within chromosomes to cause rapid changes in genomes and drastically alter phenotypes. 4. The bacterial chromosome can be mapped with great precision, using Hfr conjugation in combination with transformational and transductional mapping techniques. 5. Recombination of virus genomes occurs when two viruses with homologous chromosomes infect a host cell at the same time.
 is donated to the chromosome of a recipient cell (the endogenote) and integrated into it. 2. The actual transfer of genetic material between bacteria usually takes place in one of three ways: direct transfer between two bacteria temporarily in physical contact (conjugation), transfer of a naked DNA fragment (transformation), or transport of bacterial DNA by bacteriophages (transduction). 3. Plasmids and transposable elements can move genetic material between bacterial chromosomes and within chromosomes to cause rapid changes in genomes and drastically alter phenotypes. 4. The bacterial chromosome can be mapped with great precision, using Hfr conjugation in combination with transformational and transductional mapping techniques. 5. Recombination of virus genomes occurs when two viruses with homologous chromosomes infect a host cell at the same time.")
3
13.1 Bacterial Recombination: General Principles
1). General recombination: the most common form. usually involves a reciprocal exchange between a pair of homologous DNA sequences. it can occur anyplace on the chromosome, it results from DNA strand breakage and reunion leading to crossing-over (figure 13.2). by the products of rec genes such as the recA protein. Figure 13.2 The Holliday Model for Reciprocal General Recombination.
. General recombination: the most common form. usually involves a reciprocal exchange between a pair of homologous DNA sequences. it can occur anyplace on the chromosome, it results from DNA strand breakage and reunion leading to crossing-over (figure 13.2). by the products of rec genes such as the recA protein. Figure 13.2 The Holliday Model for Reciprocal General Recombination.")
4
1). General recombination (cont’d):
E.g., in bacterial transformation: Nonreciprocal General Recombination Figure 13.3 Nonreciprocal General Recombination. The Fox model for nonreciprocal general recombination. This mechanism has been proposed for the recombination occurring during transformation in some bacteria.

5
2) Site-specific recombination:
particularly important in the integration of virus genomes into bacterial chromosomes, the genetic material is not homologous with the chromosome, the responsible enzymes are specific for the particular virus and its host. 3) Replicative recombination: It accompanies the replication of genetic material and does not depend on sequence homology. It is used by some genetic elements that move about the chromosome.
 Replicative recombination: It accompanies the replication of genetic material and does not depend on sequence homology. It is used by some genetic elements that move about the chromosome.")
6
13.2 Bacterial Plasmids Plasmids: small double-stranded DNA molecules, usually circular, exist independently of host chromosomes, are present in many bacteria, also in some yeasts and other fungi, own replication origins and autonomously replicating and stably inherited, a replicon is a DNA molecule or sequence that has a replication origin and is capable of being replicated, plasmids have relatively few genes, generally less than 30, their genetic information is not essential to the host, single-copy plasmids produce only one copy per host cell, multicopy plasmids may be present at concentrations of 40 or more per cell.

8
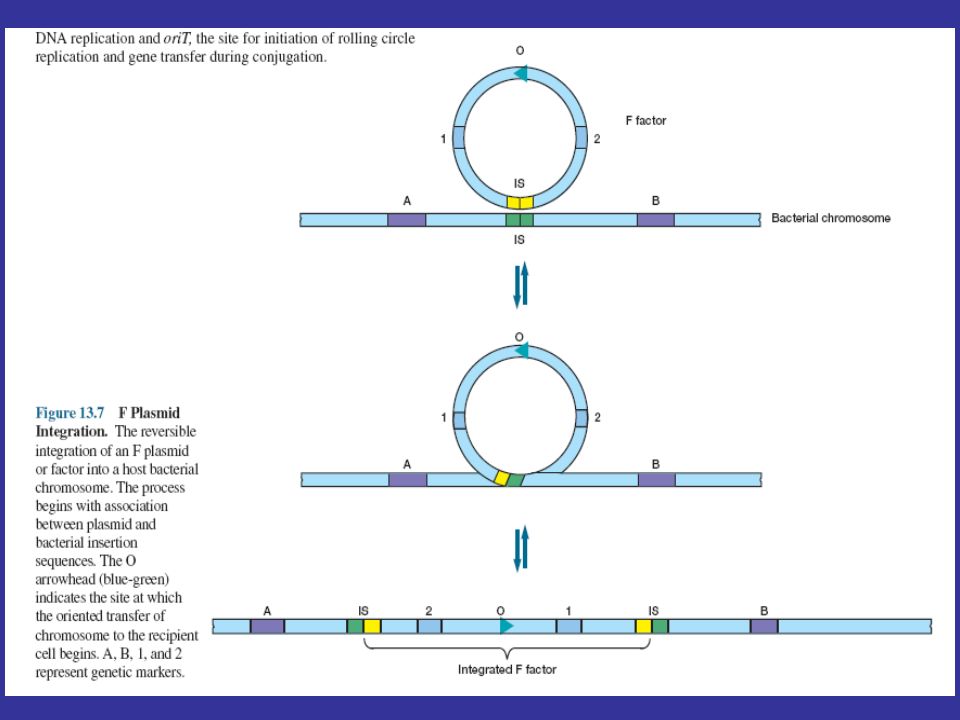
Fertility or F Factors playing a major role in conjugation in E. coli about 100 kilobases (kb) long bearing genes responsible for cell attachment and plasmid transfer between specific bacterial strains during conjugation. most of the information required for plasmid transfer is located in the tra operon, which contains at least 28 genes. many of these direct the formation of sex pili that attach the F+ cell to an F- cell. other gene products aid DNA transfer. several insertion sequences that assist plasmid integration into the host chr. an episome can exist outside the bacterial chromosome or be integrated into it.

10
Resistance Factors (R factors)
genes that code for enzymes capable of destroying or modifying antibiotics such as ampicillin, chloramphenicol, and kanamycin the resistance genes are often within a transposon usually not integrated into the host chromosome Some have only a single resistance gene, others can have as many as eight bacterial strains rapidly develop multiple resistance plasmids. many conjugative R factors can spread throughout a population. Often, nonconjugative R factors also move between bacteria during plasmid promoted conjugation. some are readily transferred between species The R factors can then be transferred to more pathogenic genera such as Salmonella or Shigella, causing even greater public health problems

11
Col Plasmids Bacteria also harbor plasmids with genes that may give them a competitive advantage in the microbial world. Bacteriocins are bacterial proteins that destroy other bacteria. They usually act only against closely related strains. Bacteriocins often kill cells by forming channels in the plasma membrane, thus increasing its permeability. They also may degrade DNA and RNA or attack peptidoglycan and weaken the cell wall. Col plasmids contain genes for the synthesis of bacteriocins known as colicins, which are directed against E. coli. Similar plasmids carry genes for bacteriocins against other species. For example, Col plasmids produce cloacins that kill Enterobacter species. Clearly the host is unaffected by the bacteriocin it produces. Some Col plasmids are conjugative and also can carry resistance genes. Other Types of Plasmids Several other important types of plasmids have been discovered. Some plasmids, called virulence plasmids, make their hosts more pathogenic because the bacterium is better able to resist host defense or to produce toxins. For example, enterotoxigenic strains of E. coli cause traveler’s diarrhea because of a plasmid that codes for an enterotoxin. Metabolic plasmids carry genes for enzymes that degrade substances such as aromatic compounds (toluene), pesticides (2,4-dichlorophenoxyacetic acid), and sugars (lactose). Metabolic plasmids even carry the genes required for some strains of Rhizobium to induce legume nodulation and carry out nitrogen fixation.
, pesticides (2,4-dichlorophenoxyacetic acid), and sugars (lactose). Metabolic plasmids even carry the genes required for some strains of Rhizobium to induce legume nodulation and carry out nitrogen fixation.")
12
13.3 Transposable Elements
Transposition: the movement of pieces of DNA that move around the genome of bacteria, viruses, and eucaryotic cells contain, or a mutation in which a chromosomal segment is transferred to a new position on the same or another chromosome Transposable elements or transposons: DNA segments that carry the genes required for this Transposition process and consequently move about chromosomes. Insertion sequences or IS elements: The simplest transposable elements. An IS element is a short sequence of DNA (around 750 to 1,600 bp in length) containing only the genes for those enzymes required for its transposition and bounded at both ends by identical or very similar sequences of nucleotides in reversed orientation known as inverted repeats. Between the inverted repeats is a gene that codes for an enzyme called transposase. Transposable elements also can contain genes other than those required for transposition (for example, antibiotic resistance or toxin genes). These elements often are called composite transposons or elements.
 containing only the genes for those enzymes required for its transposition and bounded at both ends by identical or very similar sequences of nucleotides in reversed orientation known as inverted repeats. Between the inverted repeats is a gene that codes for an enzyme called transposase. Transposable elements also can contain genes other than those required for transposition (for example, antibiotic resistance or toxin genes). These elements often are called composite transposons or elements.")
17
13.4 Bacterial Conjugation
The initial evidence for bacterial conjugation, the transfer of genetic information by direct cell to cell contact, came from an elegant experiment performed by Joshua Lederberg and Edward L. Tatum in 1946.

18
The evidence that physical contact of the cells was necessary for gene transfer was provided by Bernard Davis (1950), who constructed a U tube consisting of two pieces of curved glass tubing fused at the base to form a U shape with a fritted glass filter between the halves.
, who constructed a U tube consisting of two pieces of curved glass tubing fused at the base to form a U shape with a fritted glass filter between the halves.")
19
F+ x F- Mating In 1952 William Hayes demonstrated that the gene transfer observed by Lederberg and Tatum was polar. That is, there were definite donor (F+) and recipient (F-) strains, and gene transfer was nonreciprocal. He also found that in F+ x F- mating the progeny were only rarely changed with regard to auxotrophy (that is, bacterial genes were not often transferred), but F- strains frequently became F+. Hfr Conjugation When integrated, the F plasmid’s tra operon is still functional; the plasmid can direct the synthesis of pili, carry out rolling-circle replication, and transfer genetic material to an F- recipient cell. Such a donor is called an Hfr strain (for high frequency of recombination) because it exhibits a very high efficiency of chromosomal gene transfer in comparison with F- cells. F′Conjugation Because the F plasmid is an episome, it can leave the bacterial chromosome. Sometimes during this process the plasmid makes an error in excision and picks up a portion of the chromosomal material to form an F′plasmid.
 and recipient (F-) strains, and gene transfer was nonreciprocal. He also found that in F+ x F- mating the progeny were only rarely changed with regard to auxotrophy (that is, bacterial genes were not often transferred), but F- strains frequently became F+. Hfr Conjugation. When integrated, the F plasmid’s tra operon is still functional; the plasmid can direct the synthesis of pili, carry out rolling-circle replication, and transfer genetic material to an F- recipient cell. Such a donor is called an Hfr strain (for high frequency of recombination) because it exhibits a very high efficiency of chromosomal gene transfer in comparison with F- cells. F′Conjugation. Because the F plasmid is an episome, it can leave the bacterial chromosome. Sometimes during this process the plasmid makes an error in excision and picks up a portion of the chromosomal material to form an F′plasmid.")
21
13.5 DNA Transformation The second way in which DNA can move between bacteria is through transformation, discovered by Fred Griffith in Transformation is the uptake by a cell of a naked DNA molecule or fragment from the medium and the incorporation of this molecule into the recipient chromosome in a heritable form. In natural transformation the DNA comes from a donor bacterium. The process is random, and any portion of a genome may be transferred between bacteria.

22
13.6 Transduction Transduction is the transfer of bacterial genes by viruses. Bacterial genes are incorporated into a phage capsid because of errors made during the virus life cycle. The virus containing these genes then injects them into another bacterium, completing the transfer. Transduction may be the most common mechanism for gene exchange and recombination in bacteria. Generalized transduction occurs during the lytic cycle of virulent and temperate phages and can transfer any part of the bacterial genome. During the assembly stage, when the viral chromosomes are packaged into protein capsids, random fragments of the partially degraded bacterial chromosome also may be packaged by mistake.

23
Specialized or restricted transduction, when a prophage is induced to leave the host chromosome, excision is sometimes carried out improperly. The resulting phage genome contains portions of the bacterial chromosome (about 5 to 10% of the bacterial DNA) next to the integration site, much like the situation with F′ plasmids. A transducing phage genome usually is defective and lacks some part of its attachment site. The transducing particle will inject bacterial genes into another bacterium, even though the defective phage cannot reproduce without assistance.
 next to the integration site, much like the situation with F′ plasmids. A transducing phage genome usually is defective and lacks some part of its attachment site. The transducing particle will inject bacterial genes into another bacterium, even though the defective phage cannot reproduce without assistance..")
25
13.7 Mapping the Genome Using E. coli as an example, all three modes of gene transfer and recombination have been used in mapping. Hfr conjugation is frequently used to map the relative location of bacterial genes. This technique rests on the observation that during conjugation the linear chromosome moves from donor to recipient at a constant rate. In an interrupted mating experiment the conjugation bridge is broken and Hfr x F- mating is stopped at various intervals after the start of conjugation by mixing the culture vigorously in a blender.

27
13.8 Recombination and Genome Mapping in Viruses
Bacteriophage genomes also undergo recombination, although the process is different from that in bacteria. Because phages themselves reproduce within cells and cannot recombine directly, crossing-over must occur inside a host cell. In principle, a virus recombination experiment is easy to carry out. If bacteria are mixed with enough phages, at least two virions will infect each cell on the average and genetic recombination should be observed. Phage progeny in the resulting lysate can be checked for alternate combinations of the initial parental genotypes.

28
谢谢! Vielen dank!

Similar presentations






















 Lecture 10 Microbial Genetics (Text Chapter: 10.1-10.4; 10.6-10.14)>")
 Changes in the genetic codes 1- Intermicrobial exchange.>")



