Download presentation
Presentation is loading. Please wait.

1
Andrew Halestrap Calcium, mitochondria and reperfusion injury
A pore way to die Andrew Halestrap Department of Biochemistry and The Bristol Heart Institute

2
X ATP synthesis Cell lives Cell dies ATP breakdown
The Mitochondrial Permeability Transition A calcium induced non-specific pore Cyclosporin A X All solutes < 1500daltons Mitochondrial inner membrane Very selective permeability essential for ATP synthesis Mitochondria become leaky, swollen and uncoupled ATP synthesis Cell lives Cell dies ATP breakdown If the pore opens, not only are mitochondria unable to make ATP, they also breakdown ATP made by functional mitochondria and glycolysis. Triggered by high matrix [Ca ] 2+ Sensitised to Ca by: Low adenine nucleotides High phosphate Oxidative stress Conditions occurring when cells damaged e.g. by toxins or hypoxia

3
Proposed scheme for the mechanism of pore opening
Normal Impermeable State Ca Triggered by low [Ca 2+] Pathological Non-specific Pore Ca2+ Cytosol ADP Adenine nucleotide translocase ATP Matrix Binding increased by oxidative stress and thiol reagents. CyP binding increases sensitivity to [Ca]. ATP ADP Impermeable State Cyclophilin D Activated by thiol reagents and oxidative stress which decrease ADP/ATP binding Inhibited by [Mg 2+], low pH, adenine nucleotides and membrane potential (which increases ATP binding) Cyclosporin A Note that outer membrane proteins such as VDAC (porin), Bcl2 family members and the peripheral benzodiazipine receptor may be involved as regulatory or structural components
 Cyclosporin A. Note that outer membrane proteins such as VDAC (porin), Bcl2 family members and the peripheral benzodiazipine receptor may be involved as regulatory or structural components.")
4
Evidence for the involvement of Cyclophilin D
For all CsA analogues tested the K0.5 for inhibition of the peptidyl-prolyl cis-trans isomerase activity of CyP-D correlates with the K0.5 for inhibition of the MPTP The number of binding sites for CsA inhibition of the MPTP matches the number of binding sites for CsA inhibition of the PPIase activity of the mitochondrial matrix

5
CyP-D knockout mice show impaired permeability transition
Baines, C. P., Kaiser, R. A., Purcell, N. H., Blair, N. S., Osinska, H., Hambleton, M. A., Brunskill, E. W., Sayen, M. R., Gottlieb, R. A., Dorn, G. W., Robbins, J., and Molkentin, J. D. (2005). Loss of cyclophilin D reveals a critical role for mitochondrial permeability transition in cell death. Nature, 434: Basso, E., Fante, L., Fowlkes, J., Petronilli, V., Forte, M. A., and Bernardi, P. (2005). Properties of the permeability transition pore in mitochondria devoid of cyclophilin D. J Biol Chem, 280: Nakagawa, T., Shimizu, S., Watanabe, T., Yamaguchi, O., Otsu, K., Yamagata, H., Inohara, H., Kubo, T., and Tsujimoto, Y. (2005). Cyclophilin D-dependent mitochondrial permeability transition regulates some necrotic but not apoptotic cell death. Nature, 434: Schinzel, A. C., Takeuchi, O., Huang, Z., Fisher, J. K., Zhou, Z., Rubens, J., Hetz, C., Danial, N. N., Moskowitz, M. A., and Korsmeyer, S. J. (2005). Cyclophilin D is a component of mitochondrial permeability transition and mediates neuronal cell death after focal cerebral ischemia. Proc Natl Acad Sci U S A, 102:
. Loss of cyclophilin D reveals a critical role for mitochondrial permeability transition in cell death. Nature, 434: Basso, E., Fante, L., Fowlkes, J., Petronilli, V., Forte, M. A., and Bernardi, P. (2005). Properties of the permeability transition pore in mitochondria devoid of cyclophilin D. J Biol Chem, 280: Nakagawa, T., Shimizu, S., Watanabe, T., Yamaguchi, O., Otsu, K., Yamagata, H., Inohara, H., Kubo, T., and Tsujimoto, Y. (2005). Cyclophilin D-dependent mitochondrial permeability transition regulates some necrotic but not apoptotic cell death. Nature, 434: Schinzel, A. C., Takeuchi, O., Huang, Z., Fisher, J. K., Zhou, Z., Rubens, J., Hetz, C., Danial, N. N., Moskowitz, M. A., and Korsmeyer, S. J. (2005). Cyclophilin D is a component of mitochondrial permeability transition and mediates neuronal cell death after focal cerebral ischemia. Proc Natl Acad Sci U S A, 102:")
6
The MPTP opens in CyP-D knockout mitochondria at high calcium loading

7
.s .10 ) Rate of shrinkage (A [Ca
Inhibition of the permeability transition by CsA and SfA is overcome by high [Ca2+] 2 4 6 8 [ C a + ] m M 1 3 5 7 R t e o f S h r i n k g ( A . s - x ) Control SfA 10 20 30 40 50 60 70 Rate of shrinkage (A 520 .s ) 80 [Ca 2+ Shrinkage 1 mM SfA or 1 mM CsA
 Control. SfA Rate of shrinkage (A s .10 ) 80. [Ca. 2+ Shrinkage. 1 mM SfA. or 1 mM CsA.")
8
Proposed scheme for the mechanism of pore opening
Normal Impermeable State Ca Triggered by low [Ca 2+] Pathological Non-specific Pore Ca2+ Cytosol Ca Very high [Ca2+] ADP Adenine nucleotide translocase ATP Matrix Binding increased by oxidative stress and thiol reagents. CyP binding increases sensitivity to [Ca]. ATP ADP Impermeable State Cyclophilin D Activated by thiol reagents and oxidative stress which decrease ADP/ATP binding Inhibited by [Mg 2+], low pH, adenine nucleotides and membrane potential (which increases ATP binding) Cyclosporin A
 Cyclosporin A.")
9
The Immunosuppressive drug Sanglifehrin A is a novel inhibitor of the MPTP
This inhibitor is Sanglifehrin A. Sanglifehrin A is and has a very different structure from Cyclosporin A. In terms of its properties: Sanglifehrin A is similar to cyclosporin A in that it binds tightly to cyclophilin A. Although it has immunosuppressive properties (by blocking T-cell proliferation in response to interleukin-2), unlike CsA, the cyclophilin-Sanglifehrin complex has no effect on the calcium-dependent protein phosphatase, calcineurin which mediates the immunosuppressant activity of CsA. CLICK Cyclosporin A Sanglifehrin A Like CsA, SfA also binds tightly to CyP- A (K nM) Unlike CsA, the CyPA-SfA complex has no effect on calcineurin
, unlike CsA, the cyclophilin-Sanglifehrin complex has no effect on the calcium-dependent protein phosphatase, calcineurin which mediates the immunosuppressant activity of CsA. CLICK. Cyclosporin A. Sanglifehrin A. Like CsA, SfA also binds tightly to CyP- A (K nM) Unlike CsA, the CyPA-SfA complex has no effect on calcineurin.")
10
SfA inhibits the peptidyl prolyl cis trans isomerase activity of CyP-D and inhibits the permeability transition ) 6 1 6 - s ( 30 s A520 0.05 Control 150 nM SfA 150 nM CsA 500 nM SfA 1 mM SfA 1 mM CsA 500 nM CsA Ca2+ De-energised swelling s -1 i s 5 PPiase activity of CyP-D y 5 l o r d y 4 h 4 peptide hydrolysis (s ) Rate constant for e d t i 3 p 3 e Ki 2nM p r o f 2 2 t n a t s 1 n 1 o c e t a R 5 10 15 20 25 100 5 1 1 5 2 2 5 1 [Sanglifehrin] (nM) [ S a n g l f e h r i n ] n M
 s. ( 30 s. A Control. 150 nM SfA. 150 nM CsA. 500 nM SfA. 1 mM SfA. 1 mM CsA. 500 nM CsA. Ca2+ De-energised swelling. s. -1. i. s. 5. PPiase activity of CyP-D. y. 5. l. o. r. d. y. 4. h. 4. peptide hydrolysis (s ) Rate constant for. e. d. t. i. 3. p. 3. e. Ki 2nM. p. r. o. f t. n. a. t. s. 1. n. 1. o. c. e. t. a. R [Sanglifehrin] (nM) [ S. a. n. g. l. f. e. h. r. i. n. ] n. M.")
12
Proposed scheme for the mechanism of pore opening
Normal Impermeable State Ca Triggered by low [Ca 2+] Pathological Non-specific Pore Ca2+ Cytosol Ca Very high [Ca2+] ADP Adenine nucleotide translocase ATP Matrix Binding increased by oxidative stress and thiol reagents. CyP binding increases sensitivity to [Ca]. ATP ADP Impermeable State Cyclophilin D Activated by thiol reagents and oxidative stress which decrease ADP/ATP binding Inhibited by [Mg 2+], low pH, adenine nucleotides and membrane potential (which increases ATP binding) Sanglifehrin A SfA X Cyclosporin A
 Sanglifehrin A. SfA. X. Cyclosporin A.")
13
Evidence for the involvement of the Adenine Nucleotide Translocase

14
Inhibition of the MPT by ADP
5 1 [ C a 2 + ] m M R t e o f s h r i n k g ( A . - 4 ) D P T l B K 7 5 1 2 3 4 [ C a + ] m M R t e o f S h r i n k g ( A . s - x ) l D P
 D. P. T. l. B. K [ C. a. + ] m. M. R. t. e. o. f. S. h. r. i. n. k. g. ( A. . s. - x. ) l. D. P.")
15
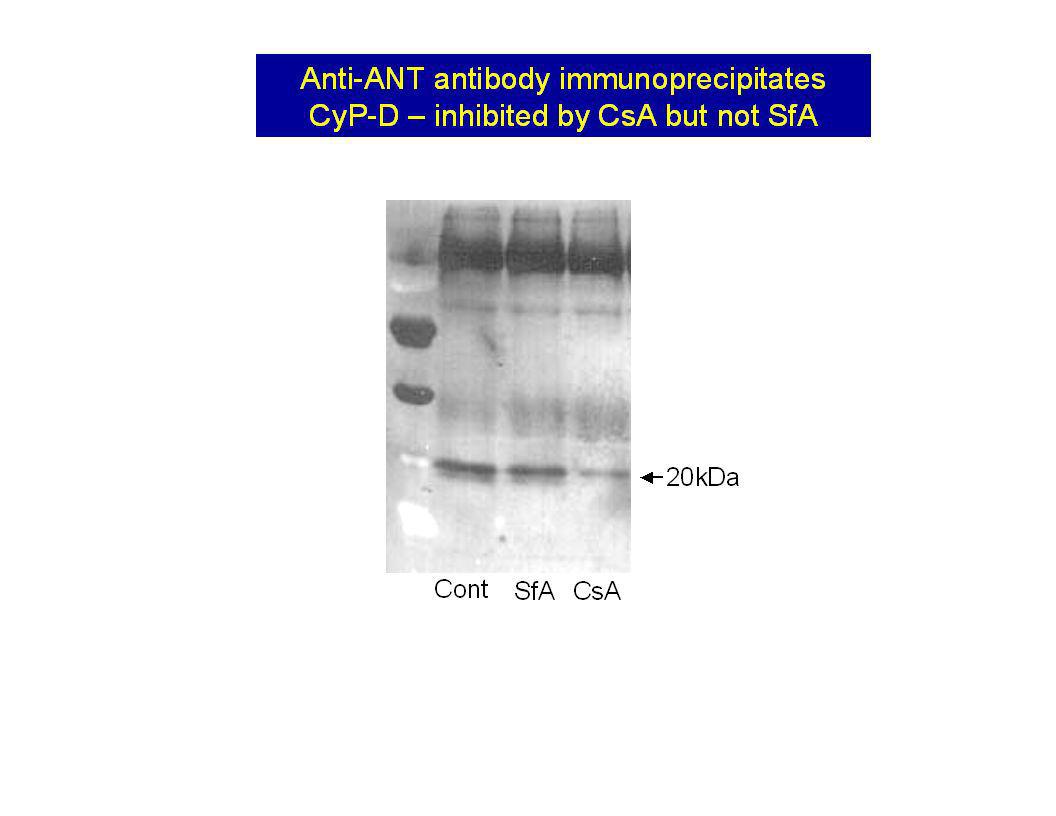
The ANT binds to immobilised CyP-D in a CsA sensitive manner
Western blots No IMM CsA + IMM CsH + IMM Plus IMM Porin Glutathione eluted proteins Cyclophilin affinity column Matrix GSH GST CyP-D Add Triton-X100-solubilised inner mitochondrial membranes Wash off non-specifically bound protein GSH Specifically bound protein 30kDa Control Diamide More ANT from diamide-treated mitochondria binds GST-CyP-D column SDS-PAGE and Western blotting with anti ANT antibodies

16
Inhibition of the MPT by ADP is antagonised by thiol modification

17
Pro Cys Cys Cys Location in the ANT of residues with potential
regulatory significance for the MPT Pro ADP Cys 61 Low N-ethylmaleimide 56 binding Cys Eosine- Cys 159 256 maleimide and high NEM CyP binding?

18
Eosine maleimide (to block Cys159) blocks ATP inhibition of the MPTP
2 4 6 8 1 [ A D P ] m M R a t e o f s h r i n k g ( 5 . - ) E l d N C + Cys159 Cys56 Cys56 plus some Cys159
 E. l. d. N. C. + Cys159. Cys56. Cys56 plus some Cys159.")
19
Pro Cys Cys Cys Location in the ANT of residues with potential
regulatory significance for the MPT Pro ADP 61 N-ethyl- maleimide Eosine- Cys 56 binding Cys Cys 159 256 Diamide and Phenylarsine oxide cross links Cys 159 to Cys 256 CyP binding?

20
Section through the carrier
Section through the carrier. At the bottom of the cone-shaped cavity, the hexapeptide (RRMMM signature) can be seen. The conical pit open to the outside and the RRR sequence spanning through the closed part of the carrier. Nature 426, (06 November 2003) Structure of mitochondrial ADP/ATP carrier in complex with carboxyatractyloside EVA PEBAY-PEYROULA1, CÉCILE DAHOUT-GONZALEZ2, RICHARD KAHN1, VÉRONIQUE TRÉZÉGUET3, GUY J.-M. LAUQUIN3 & GÉRARD BRANDOLIN2
 can be seen. The conical pit open to the outside and the RRR sequence spanning through the closed part of the carrier. Nature 426, (06 November 2003) Structure of mitochondrial ADP/ATP carrier in complex with carboxyatractyloside. EVA PEBAY-PEYROULA1, CÉCILE DAHOUT-GONZALEZ2, RICHARD KAHN1, VÉRONIQUE TRÉZÉGUET3, GUY J.-M. LAUQUIN3 & GÉRARD BRANDOLIN2.")
21
Nature 427, (29 January 2004); The ADP/ATP translocator is not essential for the mitochondrial permeability transition pore JASON E. KOKOSZKA1,4,*, KATRINA G. WAYMIRE1,4, SHAWN E. LEVY4,*, JAMES E. SLIGH4,*, JIYANG CAI5, DEAN P. JONES5, GRANT R. MACGREGOR1,2,4 & DOUGLAS C. WALLACE1,3,4 Key point: Mice with a double ANT knockout in their liver mitochondria still show a cyclosporin-sensitive MPTP. However, it is less sensitive to Ca2+ and is not blocked by ADP or activated by CAT

22
Some interesting questions
1. How can mice survive with a liver that lacks ANT? Mitochondrial export of ATP is essential for gluconeogenesis and urea synthesis for example. Do the mice have complete liver ANT knockout? 2. Even if the ANT is not functioning as the MPTP pore in the knockout, it may still do so in the normal mouse. Can other members of the mitochondrial carrier family, present in MUCH lower quantity than the ANT, act as the CyP-D binding membrane pore structure if ANT is lacking?

23
NADH NAD+ NADH OAA A OAA MAL X CyP-D ANT MPT Ca MDH MDH Pore opens A
340 OAA MAL X ANT Porin Silver stain HA HiT CyP-D ANT MPT Ca 2+ NADH NAD+ OAA MAL MDH Pore opens MDH CyP-D CyP-D + CsA 7.5 28 62 250 m M Ca 6.5 A 340 0.02 3 min CsA present in buffer CsA not present in buffer 2 min A 340 0.01 120 m M Ca 2 g/ml CyP-D 2+ Data of Jeremy Gillespie

24
Does MPTP opening occur in reperfusion injury?

25
Reversible reperfusion injury Irreversible reperfusion injury
Ischaemia (major reduction or total loss of blood flow) ATP drops and cell begins to deteriorate as a result of metabolic and ionic disturbances (e.g. lactic acid build up, low pH, high [Na] and [Ca], free radicals) Short period of ischaemia Mild to moderate damage Reversible on reperfusion Reversible reperfusion injury "Stunned" heart eventually recovers fully Long period of ischaemia Severe damage exacerbated on reperfusion which leads to cell death and necrosis Associated with swollen amorphous mitochondria that are uncoupled and have impaired respiration. They look as if the pore has opened. Irreversible reperfusion injury
 ATP drops and cell begins to deteriorate as a result of metabolic and ionic. disturbances (e.g. lactic acid build up, low pH, high [Na] and [Ca], free radicals) Short period of ischaemia. Mild to moderate damage. Reversible on reperfusion. Reversible reperfusion injury. Stunned heart eventually. recovers fully. Long period of ischaemia. Severe damage exacerbated. on reperfusion which leads to. cell death and necrosis. Associated with swollen amorphous. mitochondria that are uncoupled and. have impaired respiration. They look as if the pore has opened. Irreversible reperfusion injury.")
26
Repair of damage Cell survives Further damage Cell dies
Ischaemia / Reperfusion ATP depletion Disruption of ionic homeostasis Calcium overload Oxidative Stress Damage to intracellular components MPTP closed - High ATP Low ATP – MPTP open Repair of damage Cell survives Further damage Cell dies Mitochondria LIFE DEATH Judge and executioner

27
The "Hot DOG" technique for measuring pore opening
[ H]-DOG (2-deoxyglucose) 3 Load heart cells with 2-deoxyglucose (DOG) which is trapped in the cytosol as DOG-6-P DOG DOG-6-P DOG-6-P X Pore closed Pore open DOG-6-P only enters mitochondria if pore opens The amount of [3H]-DOG-6-P in mitochondria is used as an indicator of pore opening. Corrections are made variations in cell loading with DOG (measure whole tissue DOG) and mitochondrial recovery (citrate synthase) Isolate mitochondria in EGTA buffer. Open pores rapidly close and trap DOG+ DOG-6-P in matrix. t (min) ISCHAEMIA 20 100 50 60 125 3H-DOG-loading Isolated mitochondria
 3. Load heart cells with 2-deoxyglucose. (DOG) which is trapped in the cytosol as DOG-6-P. DOG. DOG-6-P. DOG-6-P. X. Pore closed. Pore open. DOG-6-P only enters mitochondria if pore opens. The amount of [3H]-DOG-6-P in mitochondria is used as an indicator of pore opening. Corrections are made variations in cell loading with DOG (measure whole tissue DOG) and mitochondrial recovery (citrate synthase) Isolate mitochondria in EGTA buffer. Open pores rapidly close and trap DOG+ DOG-6-P in matrix. t (min) ISCHAEMIA H-DOG-loading. Isolated mitochondria.")
28
Pore opens as pH returns to normal. (Pore is inhibited at pH<7.0)
Time dependence of mitochondrial pore opening and pH recovery during reperfusion of hearts after 30 min ischaemia pH DOG Pore opens as pH returns to normal. (Pore is inhibited at pH<7.0) 30 min ischemia PI Time of postischaemic reperfusion (min) - 0 - 100 mm Hg Ischaemia Data of Paul Kerr
 30 min ischemia. PI. Time of postischaemic reperfusion (min) mm Hg. Ischaemia. Data of Paul Kerr.")
29
Does prevention of MPTP opening protect hearts from reperfusion injury?

30
Ischaemic Preconditioning
Protecting hearts from reperfusion injury (Cyclosporin A, Sanglifehrin A and CyP-D knockout) Direct inhibitors of the MPTP Pyruvate Less oxidative stress (Free radical scavengers e.g. propofol) Low pH and [Ca2+]mito Ischaemic Preconditioning
 Direct inhibitors of the MPTP. Pyruvate. Less oxidative stress. (Free radical scavengers e.g. propofol) Low pH and [Ca2+]mito. Ischaemic Preconditioning.")
31
Cyclosporin A and Sanglifehrin A protects hearts from reperfusion injury
Data of Sam Clarke and Gavin McStay LVDP LVEDP Con SfA CsA 80 60 40 100 20 LVDP (% preischaemic value) LEVDP (mm Hg) Control SfA CsA 30 40 80 85 90 95 100 105 110 30 min Ischaemia 60 50 10 20 Time (min) * LDH release LDH released (munits/ml perfusate)
 LEVDP (mm Hg) Control. SfA. CsA min Ischaemia Time (min) * LDH release. LDH released (munits/ml perfusate)")
32
Baines et al (2005). Nature, 434: 658-662.
The hearts of CyP-D knockout mice are protected from reperfusion injury Baines et al (2005). Nature, 434: Nakagawa et al (2005).. Nature, 434:
. Nature, 434: Nakagawa et al (2005).. Nature, 434:")
33
Cyclosporin protects neurons against hypoglycaemic and ischaemic damage
Friberg, H.; FerrandDrake, M.; Bengtsson, F.; Halestrap, A. P., Wieloch, T. (1998) Cyclosporin A, but not FK 506, protects mitochondria and neurons against hypoglycemic damage and implicates the mitochondrial permeability transition in cell death. J Neurosci 18: Matsumoto, S.; Friberg, H.; FerrandDrake, M., Wieloch, T. (1999) Blockade of the mitochondrial permeability transition pore diminishes infarct size in the rat after transient middle cerebral artery occlusion. J Cerebral Blood Flow Metab 19: Toluidine blue stained Control 30 min insulin Hypoglycaemia 30 min + CsA Hypoglycaemia Control 30 min insulin Hypoglycaemia 30 min + CsA
 Cyclosporin A, but not FK 506, protects mitochondria and neurons against hypoglycemic damage and implicates the mitochondrial permeability transition in cell death. J Neurosci 18: Matsumoto, S.; Friberg, H.; FerrandDrake, M., Wieloch, T. (1999) Blockade of the mitochondrial permeability transition pore diminishes infarct size in the rat after transient middle cerebral artery occlusion. J Cerebral Blood Flow Metab 19: Toluidine blue. stained. Control. 30 min insulin. Hypoglycaemia. 30 min + CsA. Hypoglycaemia. Control. 30 min insulin. Hypoglycaemia. 30 min + CsA.")
34
Sensitivity of MPT to [Ca] in mitochondria from different regions of the brain
Cerebellum Cortex Hippocampus [Ca2+] mM 100 50 20 Rate of swelling 150 250 200 100 mM Ca 2mM EGTA Cerebellum A520 Hippocampus 1min Friberg, H.; Connern, C.; Halestrap, A. P., and Wieloch, T. (1999) J. Neurochem. 72:
![Sensitivity of MPT to [Ca] in mitochondria from different regions of the brain](http://slideplayer.com/slide/781918/3/images/34/Sensitivity+of+MPT+to+%5BCa%5D+in+mitochondria+from+different+regions+of+the+brain.jpg "Cerebellum. Cortex. Hippocampus. [Ca2+] mM Rate of swelling mM Ca. 2mM EGTA. Cerebellum. A520. Hippocampus. 1min. Friberg, H.; Connern, C.; Halestrap, A. P., and Wieloch, T. (1999) J. Neurochem. 72:")
35
Ischemia/reperfusion-mediated brain damage in WT and CypD-deficient mice
Schinzel, Anna C. et al. (2005) Proc. Natl. Acad. Sci. USA 102, Copyright ©2005 by the National Academy of Sciences
 Proc. Natl. Acad. Sci. USA 102, Copyright ©2005 by the National Academy of Sciences.")
36
Does this involve inhibition of pore opening?
Ischaemic Preconditioning Brief ischaemic episodes followed by recovery protect the heart against subsequent prolonged ischaemia and reperfusion. Does this involve inhibition of pore opening?

37
Data of Sabzali Javadov and Kelvin Lim
Protection of hearts from reperfusion injury by ischemic preconditioning ** p<0.01 Data of Sabzali Javadov and Kelvin Lim

38
Ischaemic preconditioning inhibits mitochondrial pore opening
DOG pre-loaded DOG post-loaded ** ** 15 45 70 100 130 3H-DOG-loading I I ISCHAEMIA 5 5 ISCHAEMIA 95 40 70 125 I 5 20 3H-DOG-loading 135 t (min)
")
39
Pre-ischaemia End of Ischemia 3 min Reperfusion
Ischaemic preconditioning decreases Ca2+-dependent swelling of mitochondria isolated following Ischaemia/Reperfusion 7 Control 6 IP 5 ** P<0.01 vs. Control A 4 ** 3 2 ** 1 Pre-ischaemia End of Ischemia 3 min Reperfusion Rapidly isolated mitochondria incubated under de-energised conditions in the presence of a calcium ionophore with swelling initiated by addition of 50 mM Ca2+ Data of Igor Khaliulin, Sam Clarke and Jo Parker

40
Pyruvate protects hearts from reperfusion injury
Control ischemia Pyruvate-treated ischemia 36.2 ± 9.9 Control 40 LVDP % control (6) 10 mM Pyruvate 105.3 ± 17.2 (5) (4) Post-ischemic DOG loading 57.2 ± 10.3 98.9 ± 10.8 ISCHAEMIA 100 50 60 125 3H-DOG-loading +/- 10 mM pyruvate 155 165 t (min) Post-ischaemic loading Pyruvate is: a free radical scavenger a good respiratory substrate for ATP production It causes acidification 30 (4) Mitochondrial DOG uptake (Ratio units) 20 Pre-ischaemic loading (4) +/- 10 mM pyruvate 20 50 60 100 125 10 3H-DOG-loading ISCHAEMIA t (min) No Pre-ischemic ischemia DOG loading Data of Paul Kerr
 10 mM Pyruvate ± (5) (4) Post-ischemic. DOG loading ± ± ISCHAEMIA H-DOG-loading. +/- 10 mM pyruvate t (min) Post-ischaemic loading. Pyruvate is: a free radical scavenger. a good respiratory substrate for ATP production. It causes acidification. 30. (4) Mitochondrial DOG uptake (Ratio units) 20. Pre-ischaemic loading. (4) +/- 10 mM pyruvate H-DOG-loading. ISCHAEMIA. t (min) No. Pre-ischemic. ischemia. DOG loading. Data of Paul Kerr.")
41
Effects of PROPOFOL on mitochondrial pore opening and functional recovery during reperfusion of hearts after 30 min ischaemia Control Propofol 5 10 15 20 25 MPTP opening in situ (DOG units) 40 60 80 Rate of mitochondrial swelling at 100 m M Ca 2+ DOG Swelling Rate of mitochondrial swelling at 100 mM Ca2+ LVDP EDP 70 60 50 LVDP and EDP (mm Hg) 40 30 20 10 Data of Sabzali Javadov
 Rate of mitochondrial swelling at 100. m. M Ca. 2+ DOG. Swelling. Rate of mitochondrial swelling at 100 mM Ca2+ LVDP. EDP LVDP and EDP (mm Hg) Data of Sabzali Javadov.")
43
Lactate content ATP content

44
Troponin I release Work per beat

45
The role of mitochondria in initiating the apoptotic cascade
t-Bid Bcl-2 Apoptotic signal Activation of Apaf-1 Nuclear effects Bax + - MPTP Apaf-1 Cyt-c pro-Caspase-9 dATP AIF Smac IMM OMM Mitochondrion Activation of apoptotic cascade cleavage Caspase-9 pro-Caspase-3 Caspase-3 Proteolytic - Caspase inhibitor proteins MPTP opening causes swelling and release of intermembrane proteins. Could MPTP opening be involved in apoptosis?

46
Stress to cell - ? bcl-2 APOPTOSIS NECROSIS
Mitochondrial Permeability Transition Mitochondrial swelling and outer membrane rupture Release of cytochrome c, AIF and Smac/Diablo Activation of caspases APOPTOSIS MPT pores close ATP production maintained Moderate insult pores reseal The extent of the permeability transition may determine whether cell death is necrotic or apoptotic following ischaemia and reperfusion NECROSIS MPT pores stay open ATP is depleted Severe insult pores remain open

47
? - - Role of the Mitochondrial Permeability Transition in
Apoptosis and Necrosis bcl-2 - Removal of growth factors Cytokines e.g. TNF, Fas Activation of Caspase 8 ? t-Bid and Bax migrate to mitochondrial outer membrane Multiple and interacting signalling pathways Stresses e.g. reperfusion or toxins Mitochondrial Permeability Transition Swelling and outer membrane rupture Transient opening - ? bcl-2 Release of cytochrome c, Activation of Activation of Smac/Diablo and AIF caspase 3 caspase 9 Prolonged Apoptosis requires ATP levels to be maintained, opening Induction of whereas in necrosis ATP levels fall. Transient apoptotic cascade MPT opening allows some swelling of mitochondria and cytochrome c release, but on resealing ATP levels can be restored allowing apoptosis to occur. APOPTOSIS NECROSIS

48
MRC BBSRC Wellcome Trust

Similar presentations














 Oxidative phosphorylation. Proton Motive Force ( p ) PMF is the energy of the proton concentration gradient The chemical ( pH=>")




