Download presentation
Presentation is loading. Please wait.

1
Renal Physiology II Urination Tubular Transport Countercurrent

2
Getting Urine from the kidney to the outside. (Urination or micturition) Processed tubular fluid is dumped by the collecting system into the renal pelvis where it enters the ureters. Ureters: conduits that propel urine by peristaltic contractions toward the bladder. Bladder: a muscular “bag” that holds urine and forces it by contration. Urethra: the conduit for urine from the baldder to the outside
 Processed tubular fluid is dumped by the collecting system into the renal pelvis where it enters the ureters. Ureters: conduits that propel urine by peristaltic contractions toward the bladder. Bladder: a muscular bag that holds urine and forces it by contration. Urethra: the conduit for urine from the baldder to the outside.")
3
Urethra Pelvis: Collects urine from collecting ducts. In the pelvis there are “electrical pacemaker” cells that initiate peristaltic waves in the (2) smooth muscle sheaths of the ureteral wall. (The pelvis to ureter is a functional syncitium, not unlike the muscular wall of the heart). The frequency of the waves is 2-6/min. The pacemaker cells seem to be stimulated by the stretch of urine filling the pelvis. The movement of the peristaltic wave is about 2-6 cm/sec., traveling from its origin at the pelvis down to the bladder.
 smooth muscle sheaths of the ureteral wall. (The pelvis to ureter is a functional syncitium, not unlike the muscular wall of the heart). The frequency of the waves is 2-6/min. The pacemaker cells seem to be stimulated by the stretch of urine filling the pelvis. The movement of the peristaltic wave is about 2-6 cm/sec., traveling from its origin at the pelvis down to the bladder..")
4
Page 752 Anatomy of the bladder and ureter. On the right is the electrical profile of a peristaltic wave passing down the muscular wall of the ureter.

5
The peristaltic waves propel the urine along the ureter, generating a pressure head of which changes from a baseline of 2-5 cm H2O up to 20-80 cm H2O. While peristalysis is independent of nerve input, the action of symapthetic nerves innervating the ureter may modify the rate or force of peristalsis. Interruption of the flow of urine by an obstruction (such as a kidney stone) stops flow, increases pressure which can back up through the ureter into the pelvis, and increase the nephron and subcapsular hydrostatic pressure. This may result in the condition hydronephrosis in which the medulla is damaged and may even be sloughed off, leaving a hollow kidney. Obviously this condition impares the concentrating ability fo the kidney. There are autonomic pain fibers in the ureter which account for the acute pain when a kidney stone is formed.
 stops flow, increases pressure which can back up through the ureter into the pelvis, and increase the nephron and subcapsular hydrostatic pressure. This may result in the condition hydronephrosis in which the medulla is damaged and may even be sloughed off, leaving a hollow kidney. Obviously this condition impares the concentrating ability fo the kidney. There are autonomic pain fibers in the ureter which account for the acute pain when a kidney stone is formed..")
6
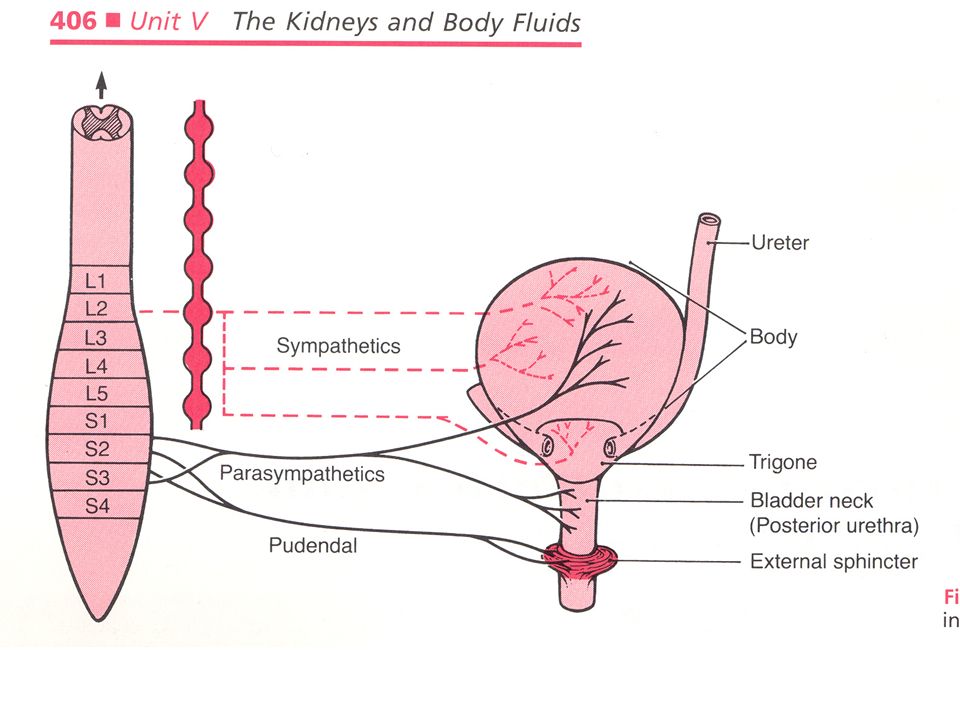
The bladder and its sphincters is also innervated with sympathetic, parasympathetic and somatic (voluntary) nerves. The wall of the bladder is composed of three muscular layers, called the “detrusor muscle.” A triangular membrane called the “trigone” acts as a valve system along with the internal sphinctors of the muscular wall to prevent urine reflux into the ureters.

7
Again, the anatomy of the bladder: note trigone. Page 752

8
Sympathetic nerves originate from the neurons ot the intermediallateral cell column from T-10 to L-2. they innervate the body of the bladder and the trigone. Parasympathetic nerves originate from S2-S4 of the sacral spinal cord. They innervate the body and neck of the bladder. Somatic innervation (voluntary or pudendal) originates from the motor neurons arising from S-2 to S-4. They innervate and control the voluntary muscles of the external sphinctor.
 originates from the motor neurons arising from S-2 to S-4. They innervate and control the voluntary muscles of the external sphinctor..")
10
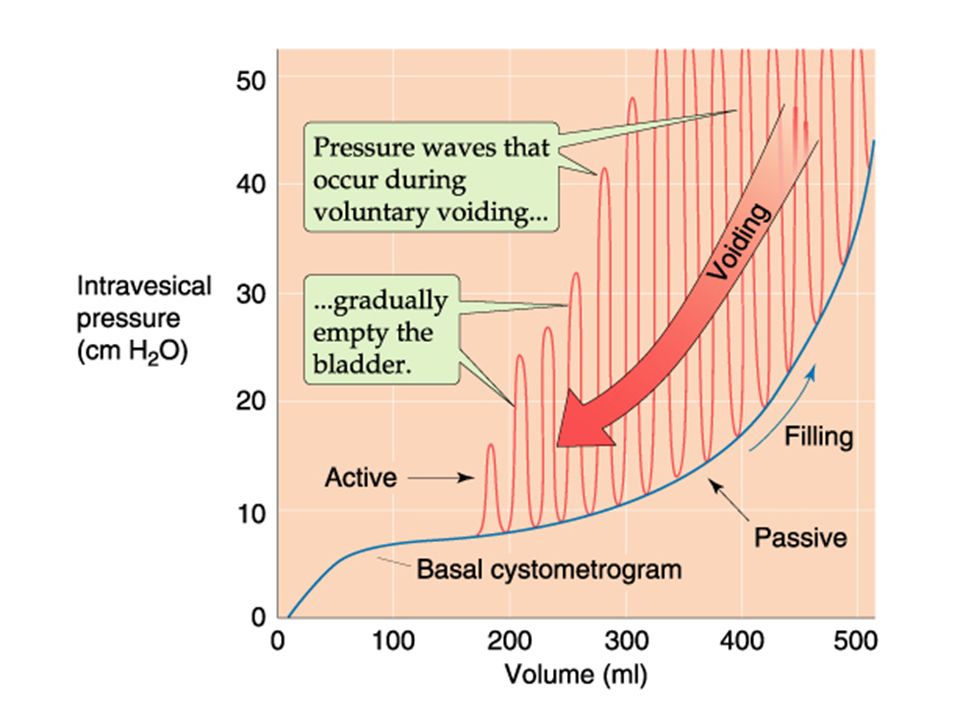
Bladder tone is derived from the volume and pressure exerted on the inside of the bladder (intervesical pressure). Increasing bladder volume by 50 ml increases pressure. As volume inceases further, the intervesical pressure increases, but not much until you get above 300 ml. then the pressure rises steeply with additional volume. (see next slide-blue line). This increase in volume and pressure increases bladder “tone” triggering the mictiurition reflex (open the flood- gates!) Efferent impulses from the brain supress the reflex (a learned reflex) until a decision is made to relax the external sphinctor using voluntary nerves. Voiding begins with relaxation of the external sphinctor, then the internal sphinctor.
. This increase in volume and pressure increases bladder tone triggering the mictiurition reflex (open the flood- gates!) Efferent impulses from the brain supress the reflex (a learned reflex) until a decision is made to relax the external sphinctor using voluntary nerves. Voiding begins with relaxation of the external sphinctor, then the internal sphinctor..")
12
Next, the detrussor muscle of the bladder wall contracts in waves (see red lines in previous figure) to expell the urine. Voluntary contraction of the abdonimal muscles further contracts the bladder, increasing the voiding. Once the bladder is empty, we are back down to the “no tone” phase (in the lower left corner of figure 32- 14) and the sphinctors can close again. The process is sterile until it leaves the body. However, because of all the organic and waste material, once out is it a good culture media.
 and the sphinctors can close again. The process is sterile until it leaves the body. However, because of all the organic and waste material, once out is it a good culture media..")
13
The urethara in the man passes through the prostate gland. As the prostate enlarges, due to age related hypertrophy or prostate cancer, it constricts the ureter and makes voiding difficult, despite intact bladder and sphincter reflexes. The purpose of the prostate is to provide urologists with a large source of income, as well as extensive extramural research funding and publications.

14
SECTION II Transport along the nephron :

15
Transport of Sodium (Na + ) and Chloride (Cl - ) Chapter 34 The filtered load of Na+ is the product of the glomerular filtration rate (GFR, 180 liters/day) and the plasma Na+ concentration (142 mM), or approximately 25,500 mM/day (equivalent to the Na+ in approximately 1.5 kg of table salt, more than nine times the total quantity of Na+ present in the body fluids. With a typical Western diet consuming approximately 120 mM of Na+ per day, the kidneys reabsorb approximately 99.6% of the filtered Na+ by the time the tubule fluid reaches the renal pelvis. Therefore, even minute variations in the fractional reabsorptive rate could lead to changes in total-body Na+ that markedly alter ECF volume, sodium balance, blood pressure, body weight and many other

16
The filtered load of Na is 25,500 mM/day, but the intake is only 120 mM/day and the output is 100 mM/day excreted plus about 20 mM in the feces and sweat. Thus, the intake equals output, so the body is in sodium balance.

17
Reabsorption of filtered Na load along the each nephron segment. Yellow boxes are the amount of filtered which is reabsorbed. Green boxes represent the amount of filtered which remains in each portion of the nephron. Pg 776

18
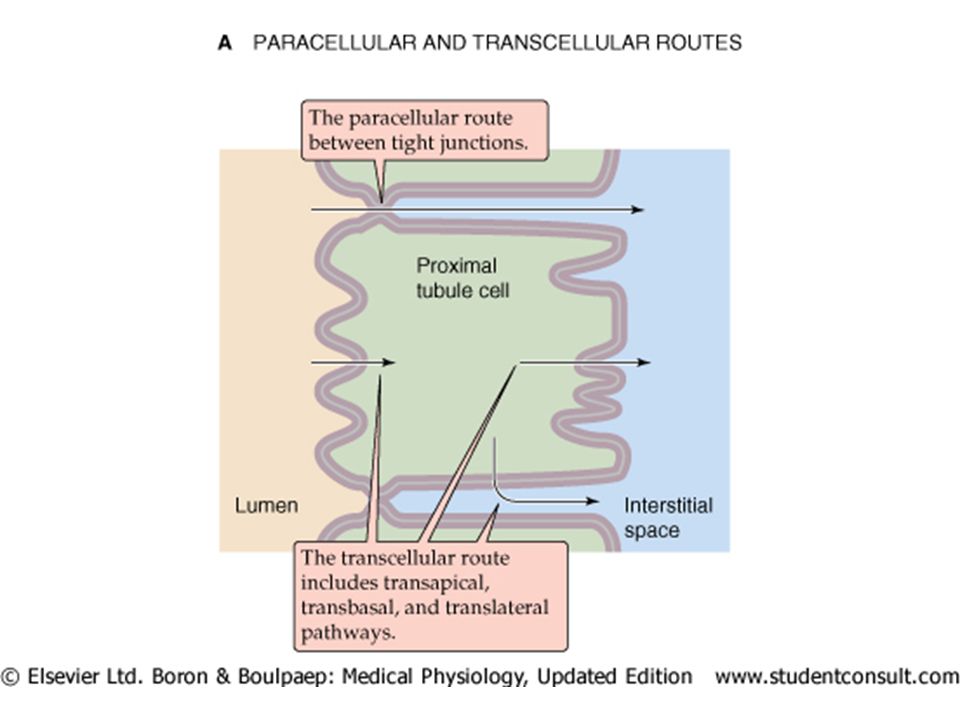
Transport of ions, and particularly of sodium from the lumen to the blood across the tubular wall is through two pathways; transcellular and paracellular. Transport is driven by two general mechanisms; 1)active transport in an energy (ATP) utilizing fashion where ions are pumped against their electrochemical gradient (“uphill”), and 2) passively down their electrochemical gradient (“downhill”) along the gradients created by the active transport.
active transport in an energy (ATP) utilizing fashion where ions are pumped against their electrochemical gradient ( uphill ), and 2) passively down their electrochemical gradient ( downhill ) along the gradients created by the active transport..")
19
Na Tubular lumen Apical Membrane Basal-lateral membrane Interstitial space Capillary Lumen Na Tubular epithelial wall Capillary wall Net pathway for sodium (Na) reabsorption from tubular lumen to capillary Start here with Na delivery End here with sodium reabsorbed and recovered
 reabsorption from tubular lumen to capillary Start here with Na delivery End here with sodium reabsorbed and recovered")
21
Para-cellular movement incorporates The transepithelial electrochemical Na gradient drives passive Na reabsorption in the proximal and thick ascending limb of the nephron. Not so for later nephron segments where the net (passive) force favors movement into the lumen. Na can also move passively in the proximal tubule (without active transport) via “solvent drag” where the movement of water (driven by active Na transport) sweeps additional Na and Cl along with it (a sort of mass-action) out of the lumen into the lateral intracellular space. The leakiness of the nephron (facillitating passive reabsorption) is greatest in the proximal, and decreases along the nephron to the papillary collecting ducts.
 force favors movement into the lumen. Na can also move passively in the proximal tubule (without active transport) via solvent drag where the movement of water (driven by active Na transport) sweeps additional Na and Cl along with it (a sort of mass-action) out of the lumen into the lateral intracellular space. The leakiness of the nephron (facillitating passive reabsorption) is greatest in the proximal, and decreases along the nephron to the papillary collecting ducts..")
22
Trans-cellular movement incorporates 1) passive entry from the lumen via the “apical” membrane into the cell down an electro-chemical gradient. The proximal, TAL and DCT use various co-transporters and exchangers, while in the collecting ducts Na enters via Na channels. 2) Active extrusion of Na out the basal-lateral membrane via a Na-K + pump which maintains intracellular Na low and K high. This exchange keeps the voltage at 70 mV (cell interior negative vs interstitium, or lumen) depending on pump activity and the voltage gradient it creates.
 Active extrusion of Na out the basal-lateral membrane via a Na-K + pump which maintains intracellular Na low and K high. This exchange keeps the voltage at 70 mV (cell interior negative vs interstitium, or lumen) depending on pump activity and the voltage gradient it creates..")
23
“Downhill” refers to a passive flow along an electrochemical gradient not requiring active transport. There is a net driving force due to the active pump forcing Na into the interstitium, but a net negative change favoring the lumen to draw Na back via extracellular junctions.

24
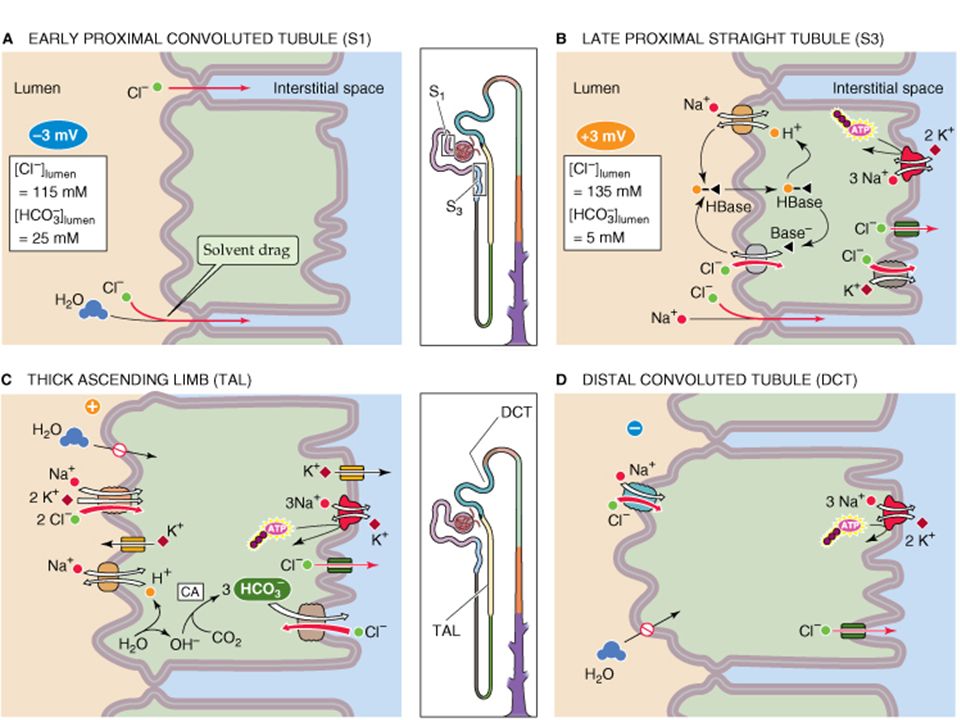
Proximal: the Na-K pump on the apical (interstitial side) membrane is the driving force for the electrochemical gradient which drives passive transport into the cell and keeps intracellular Na low, pumping against the gradient into the basal-lateral space. Passive entry into the cell is by diffusion, facillitated diffusion through a transporter or co-transporter, and by electroneutral “exchange” with hydrogen ions (H+). Na + pump Tubular lumen Interstitial space
. Na + pump Tubular lumen Interstitial space.")
25
Na+ reabsorption in different nephron segments

26
The sodium movement across the thin limbs (decending and ascending limbs) of the loop of Henle are virtually entirely passive down its electrochemical gradient and paracellular. Keep this in mind when we return to the countercurrent system!.

27
Thick Ascending Limb (TAL) of the loop of Henle. Transcellular Na reabsorption includes 1)the Na/K/2Cl co-transporter (NKCC2) which couples inward movement of these three ions in an electroneutral (2+ &2-) process driven by the downhill gradient of Na and Cl into the cell. (Note that this pump is the target of loop diuretics). Much of the K+ entering the cell is extruded via K channels down its own gradient. 2)The Na + -H + exchanger exchanging sodium for hydrogen in an electroneutral process.
the Na/K/2Cl co-transporter (NKCC2) which couples inward movement of these three ions in an electroneutral (2+ &2-) process driven by the downhill gradient of Na and Cl into the cell. (Note that this pump is the target of loop diuretics). Much of the K+ entering the cell is extruded via K channels down its own gradient. 2)The Na + -H + exchanger exchanging sodium for hydrogen in an electroneutral process..")
28
Na+ reabsorption in different nephron segments

29
Paracellular sodium transport by the thick ascending limb (TAL) (also known as the “diluting” segment). Because the lumen of the TAL is positive voltage due to the high density of K+ channels in the apical membrane, unlike all other nephron segment epithelia. This lumen-positive voltage drives sodium (and other positively charged ions) out of the lumen across the tight junctions between the cells. This paracellular pathway accounts for about half of the sodium movement out of the lumen to the basalateral spaces and the interstitium. The TAL has low water permeability, so removal of ions without water following leaves the lumen dilute (hypoosmotic) and the interstitium concentrated (hyperosmotic).
 out of the lumen across the tight junctions between the cells. This paracellular pathway accounts for about half of the sodium movement out of the lumen to the basalateral spaces and the interstitium. The TAL has low water permeability, so removal of ions without water following leaves the lumen dilute (hypoosmotic) and the interstitium concentrated (hyperosmotic)..")
30
Transcellular and paracellular Na reabsorption in different nephron segments

31
Distal Convoluted Tubule (DCT) Sodium reabsorption in the distal tubule is almost entriely due to transcellular transport. Electroneutral passive apical Na entry is due to an Na/Cl cotransporter (NCC). Unlike the NKCC2, this is independent of K+ (this pump is the target of the “thiazide” diuretics, which tqarget sodium without wasting potassium) The net movement of transcellular sodium in the DCT is driven by an ATP-utilizing basal-lateral Na + - K + pump
. Unlike the NKCC2, this is independent of K+ (this pump is the target of the thiazide diuretics, which tqarget sodium without wasting potassium) The net movement of transcellular sodium in the DCT is driven by an ATP-utilizing basal-lateral Na + - K + pump.")
32
Transcellular and paracellular Na reabsorption in different nephron segments

33
Sodium transport in the Collecting Tubules: The relatively modest Sodium reabsorption in the collecting tubules is entirely transcellular via the “principal cells.” Na enters the apical membrane via a “voltage-gated sodium channel” or “ENaC.” The basolateral movement of sodium out of the cell is driven by an energy requiring Na-K pump which establishes the gradient driving apical sodium entry. The movement of Na+ out of the lumen into the cell makes the lumen negatively charged, and the movement of K out of the cell, primarily into the basolateral interstitium makes the cell negative, for a net transepithelial voltage of -40 mV. The hormones aldosterone and vasopressin can change this site of transport.

34
Transcellular and paracellular Na reabsorption in different nephron segments

35
Medullary Collecting Duct: The inner and outer medulalry collecting ducts reabsorb only a small amount of sodium (3% of filtered load) and this is probably via ENaC on the apical membrane and the Na-K pump driving Na movement on the basal lateral membrane.
 and this is probably via ENaC on the apical membrane and the Na-K pump driving Na movement on the basal lateral membrane.")
36
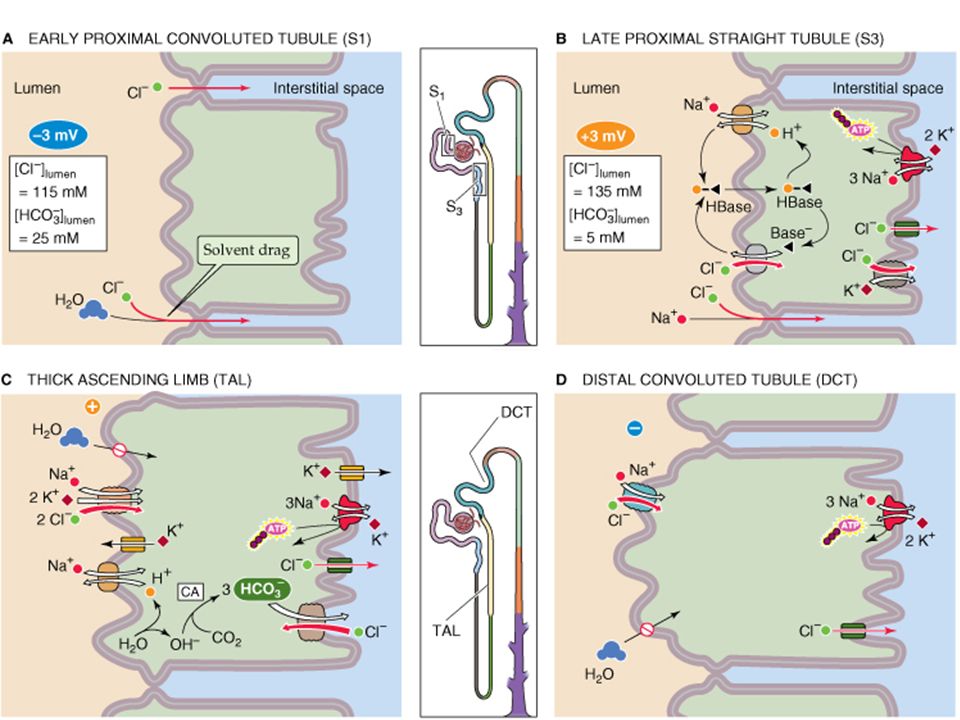
Cl - transport and reabsorption: Most Cl follows along with Na reabsorption, but the exact nature fo the movement is somewhat different. In the Proximal tubule: Early proximal tubule Cl reabsorption is mostly paracellular via solvent drag driven by the lumen negative potential. However, in the late proximal it is reabsorbed by a predominantly by transcellular pathway, driven by apical H+ exchange and active transport with Na and a K cotransporter. The lumen becomes positive actually retarding Cl reabsorption.

38
Changes in proximal TF:P ratio along the length of the proximal tubule. Note that 65% of the water is lost so inulin continues to concentrate, while osmolality and [Na} are unchanged.

39
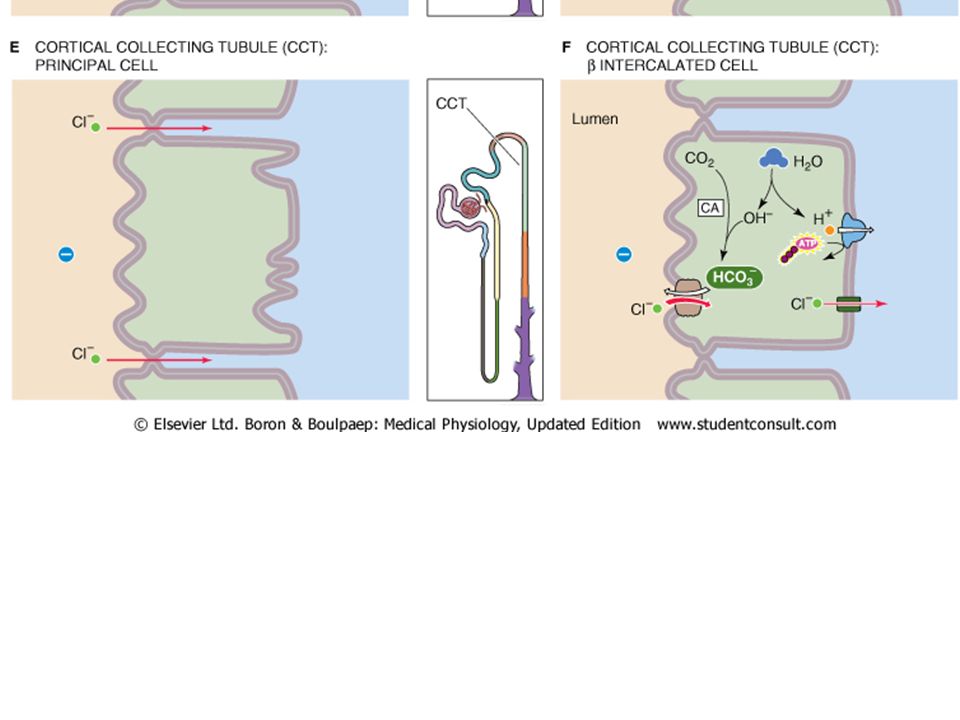
Thick ascending limb (TAL): Cl is primarly reabsorbed by the NaK2Cl co-transsporteracross the apical membrane, and basal lateral Cl channels along with active transport of sodium drive Cl into the interstieium. Distal Tubule: Apical Cl reabsorption occurs via a Na/Cl ccotransporter and is driven by Cl following Na active extrusion via a Na/K pump. Collecting ducts: The principal cell has an electrogenic pump that creates a -40 mV lumen negative potential that drives Cl- out of the lumen via paracellular routes. However, the other cell type (the intercallated cell) drives transcellular Cl movement powered by a basalateral H+ pump.
 drives transcellular Cl movement powered by a basalateral H+ pump..")
42
Water Reabsorption: In the proximal tubule, water follows sodium passibvely and isosmotically because the proximal tubule is very permeable to water. Water moves both transcellularly and paracellularly. The transcellular movement is facillitated by “aquaporin” water channels in both the apical and basalateral membranes.

43
Water reabsorption in the Thick Ascending Limb and Distal nephrons. All the distal nephrons, from TAL on, have a very low water permeability (in the presence of vasopressin). This low water permeability will be very important in understanding “countercurrent” concentration of the urine (coming later) The combination of Na transport with low water permeability produce a dilute tubular fluid with low Na and low osmolality. This facillitates later passive water reabsorption down a concentration gradient out of the nephron and into the capillary blood.
. This low water permeability will be very important in understanding countercurrent concentration of the urine (coming later) The combination of Na transport with low water permeability produce a dilute tubular fluid with low Na and low osmolality. This facillitates later passive water reabsorption down a concentration gradient out of the nephron and into the capillary blood..")
44
End here for now

45
Because of the high energy involved in active reabsorption by the nephron using ATP and the generation of ATP by oxidative metabolism, the O 2 consumption by the kidney (cortex) is very high. While the kidneys are only 0.5% of the total body weight, they account for 7-10% of O 2 consumption. The figure shows that there is a dirct correlation between sodium reabsorption and O2 consumption.

46
Regulation of Na transport by glomerulotubular balance: Changes in renal hemodynamics change Na load presented to the kidney and the “glomerulotubular balance” in the proximal tubule. When the Na load is changes, proximal reabsorption changes proportionately such that the amount reabsorbed is a constant fraction of that filtered. Absolute reabsorption of Na (mM/min)
.")
47
What factors regulate glomulotubular (G-T) balance? Starling forces are altered when the filtration fraction is changed. Normally in the capillaries, hydrostatic pressure is low and oncotic pressure high. This may be changed by altering the amount of solutes and water filtered. Thus, the changes that take place reflect the changes in filtered load and what the proximal must deal with. Internally the presentation of more Na into the lumen means more “substrate” to be reabsorbed. Altered filtered load will also change the Starling forces in the capillary and affect the net driving force for reabsorption. Finally, certain “humoral factors” (angiotensin, nitric oxide) directly affect Na reabsorption and so can alter G-T balance
 directly affect Na reabsorption and so can alter G-T balance.")
48
Humoral factors modulate glomerulotubular balance Renin-angiotensin-aldosterone: aldosterone stimulates Na reabsorption in the collecting tubules via ENaC channels and the basal lateral Na-K pump. Angiotensin itself can directly reduce Na reabsorption. Sympathetic renal innervation release norepinephrine release to reduce RBF, GFR, and reduce Na excretion. Renal nerves also may stimulate renin and further induce the renin angiotensin system. Arginine vasopressin (ADH) from the posterior pituitary which produces a concentrated urine and conserves H 2 O. It acts by increasing the water permeability of the collecting tubules and increasing Na reabsorption.
 from the posterior pituitary which produces a concentrated urine and conserves H 2 O. It acts by increasing the water permeability of the collecting tubules and increasing Na reabsorption..")
49
Distal Flow-dependent reabsorption Distal sodium reabsorption also increases with an increased distal sodium load. Increased flow and Na delivery increases the work load of the pumps which can work more at higher flows. However, this is not a linear relationship as in the proximal, as the cululative Na reabsorption may only increase at a ration of half that of the increase in distal delivery.

50
NEXT SECTION Transport of other molecules: Urea, glucose, Phosphate, Calcium, magnesium and organic solutes. Beyond sodium, chloride and water, the kidney handles other substances which are important in homeostasis.

51
Urea: is produced primarly in the liver as a bi- product of amino acid metabolism. The MAIN pathway for the body to eliminate this “waste product” is through the kidney. Urea is freely filtered by the kidney, then both secreted and reabsorbed (overall, there is net reabsorption, so the clearance of urea is 60% less than the GFR). Proximal tubule and medullary collecting duct are the main sites for reabsorption. The thin limbs of the loop of Henle are the primary sites for secretion.
. Proximal tubule and medullary collecting duct are the main sites for reabsorption. The thin limbs of the loop of Henle are the primary sites for secretion..")
52
Urea

53
Water leaves the proximal tubular lumen dragging urea, so reabsorption is linked to flow. However, urea does not leave as efficiently as sodium, so the luminal urea concentration rises, creating a diffusion gradient out of the tubule. In the loop of Henle, in the medullary interstitium, urea concentration is high. The thin limbs of the loop have a urea transporter (UT2) which secretes urea into the lumen via facillitated diffusion. Luminal concentration rises to 110% of the filtered load by the end of the loop. In the medullary collecting duct, urea is reabsorbed by apical and basalateral facillitated transport (UT) due to the high luminal urea concentration. This movement into the deep medulla helps keep interstitial urea concentration high for the countercurrent system.
 which secretes urea into the lumen via facillitated diffusion. Luminal concentration rises to 110% of the filtered load by the end of the loop. In the medullary collecting duct, urea is reabsorbed by apical and basalateral facillitated transport (UT) due to the high luminal urea concentration. This movement into the deep medulla helps keep interstitial urea concentration high for the countercurrent system..")
54
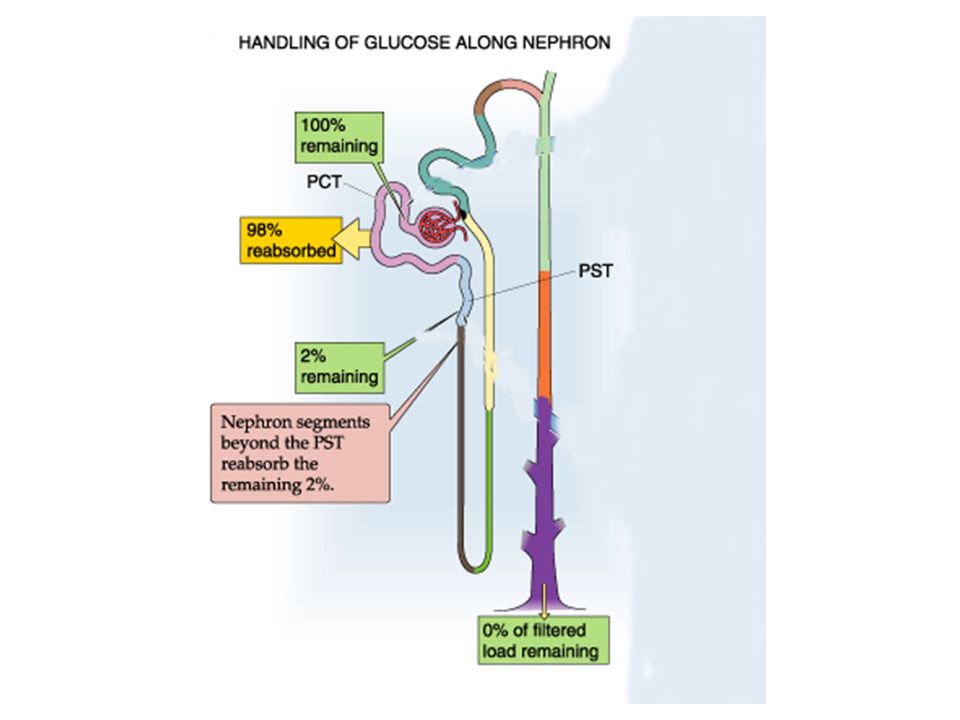
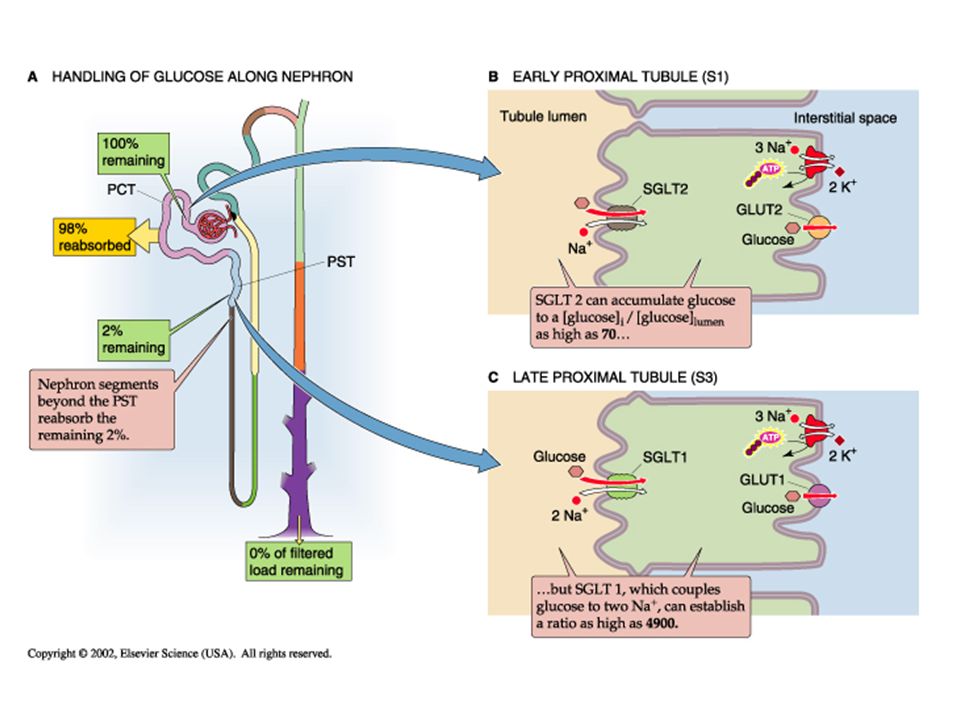
Glucose Transport: Glucose in the blood is tightly regulated by insulin at 4-5 mM (70-100 mg/dL). Glucose is freely filtered by the glomerulus, by 98% of the filtered glucose is reabsorbed by the proximal tubule. Normally, virtually no glucose appears in the urine. Proximal glucose transport is active coupled to sodium transport via sodium-glucose co-transporters (SGLT) on the apical membrane and across the basalateral membrane by facillitated diffusion (via GLUT 1 and 2). In the early proximal it is driven by a high capacity, low affinity transporter (SGLT2), but later along the proximal by a low capacity, high affinity transporter (SGLT1).
 on the apical membrane and across the basalateral membrane by facillitated diffusion (via GLUT 1 and 2). In the early proximal it is driven by a high capacity, low affinity transporter (SGLT2), but later along the proximal by a low capacity, high affinity transporter (SGLT1)..")
57
While glucose is normaly totally reabsorbed, if the plasma levels start to increase, the filtered load will increase and it can surpass the ability of the nephron to totally reabsorb it. This maximum level is referred to as the “transport maximum (T m ).” It reflects a point where the SGLT transporters are totally saturated, so any filtered glucose beyond that maximal amount reabsorbed will begin appearing in the urine. Coupled to the saturation of the transporters, the clearance of glucose (which is normally zero) will begin to rise, reflecting a fraction of the total filtered which now appears in the urine. See figure 35-5 on page 794
. It reflects a point where the SGLT transporters are totally saturated, so any filtered glucose beyond that maximal amount reabsorbed will begin appearing in the urine. Coupled to the saturation of the transporters, the clearance of glucose (which is normally zero) will begin to rise, reflecting a fraction of the total filtered which now appears in the urine. See figure 35-5 on page 794.")
58
Glucose filtration (yellow), reabsorption (red) and excretion (green) vary as the transport maximum of the nephron is reached at around 250 mg/dL. Glucose clearance is normally 0, but beyond the capacity of the nephron to reabsorb, it starts increasing as a percentage of the filtered load spills over into the urine.

59
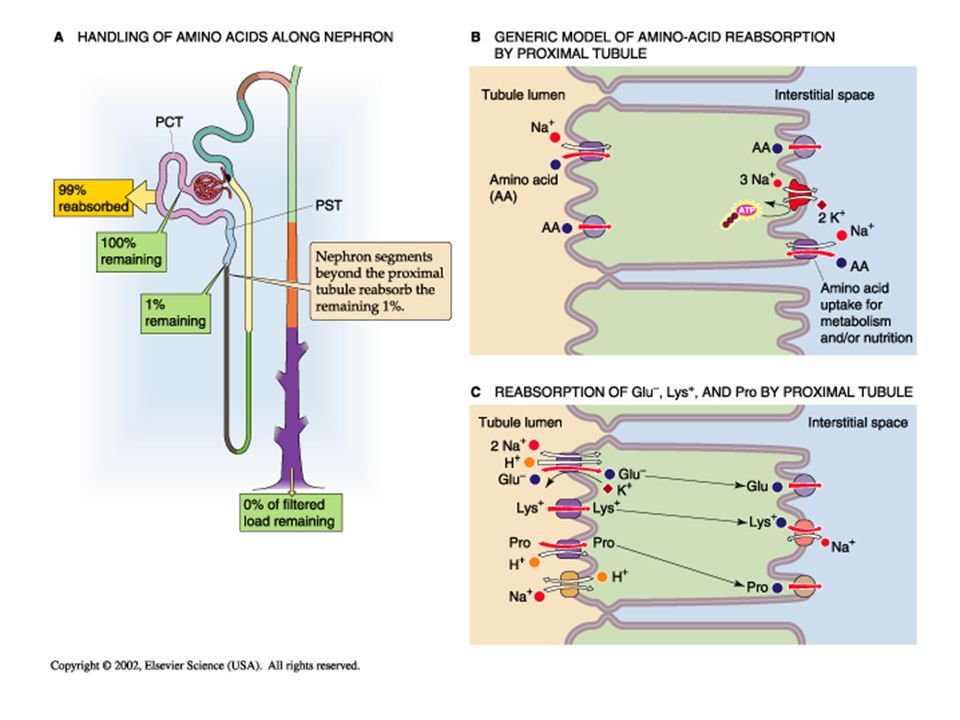
Reabsorption of Amino Acids; The body does not want to lose their nutrient substrate amino acids, so while the kidney freely filters them, they are normally totally reabsorbed. Apical amino acid transport is typically active by a sodium- dependent co-transporter, though some amino acids are reabsorbed via Na-independent facilitated diffusion. On the basal lateral membrane most amino acids exit the cell by facilitated diffusion. In some cases, due to similar molecular structures, the amino acids may exhibit competitive inhibition of transport. Amino acid transport kinetics are similar to glucose, in that they exhibit a transport maximum and may saturate if plasma levels are too high.

61
Reabsorption of other molecules by the kidney. Oligopeptides: filtered oligopeptides are totally reabsorbed in the proximal tubule. Proteins: proteins (and protein fragments) are typically not filtered, but those that are get reabsorbed by receptor-mediated endocytosis, metabolized and taken back into the blood stream, so that only trace amounts show up in the urine. Carboxylates (pyruvate and lactate) are products of anaerobic glucose matabolism and are sued as intermediates in the citric acid cycle. These are totally conserved by reabsorption by the kidney Organic anions and cations are metabolic products secreted by the late proximal tubule. Phosphate: Similar to calcium, 90% of filtered ionic free phosphate is reabsorbed by the kidney, mostly in the proximal.
 are typically not filtered, but those that are get reabsorbed by receptor-mediated endocytosis, metabolized and taken back into the blood stream, so that only trace amounts show up in the urine. Carboxylates (pyruvate and lactate) are products of anaerobic glucose matabolism and are sued as intermediates in the citric acid cycle. These are totally conserved by reabsorption by the kidney Organic anions and cations are metabolic products secreted by the late proximal tubule. Phosphate: Similar to calcium, 90% of filtered ionic free phosphate is reabsorbed by the kidney, mostly in the proximal..")
62
Calcium

63
Calcium in the plasma: The total concentration of calcium in plasma is normally 2.2 to 2.7 mM (8.8-10.6 mg/dl). Some 40% binds to plasma proteins, mainly albumin, and constitutes the nonfilterable fraction. The filterable portion, approximately 60% of total plasma calcium, consists of two moieties. 1)approximately 15% of the total, complexes with small anions such as carbonate, citrate, phosphate, and sulfate. 2)approximately 45% of total calcium, is the ionized Ca 2+ that one may measure with Ca 2+ -sensitive electrodes or dyes. It is the concentration of this free, ionized calcium that the body tightly regulates; plasma [Ca 2+ ] normally is 1.0 to 1.3 mM (4.0-5.2 mg/dl).
approximately 15% of the total, complexes with small anions such as carbonate, citrate, phosphate, and sulfate. 2)approximately 45% of total calcium, is the ionized Ca 2+ that one may measure with Ca 2+ -sensitive electrodes or dyes. It is the concentration of this free, ionized calcium that the body tightly regulates; plasma [Ca 2+ ] normally is 1.0 to 1.3 mM ( mg/dl)..")
65
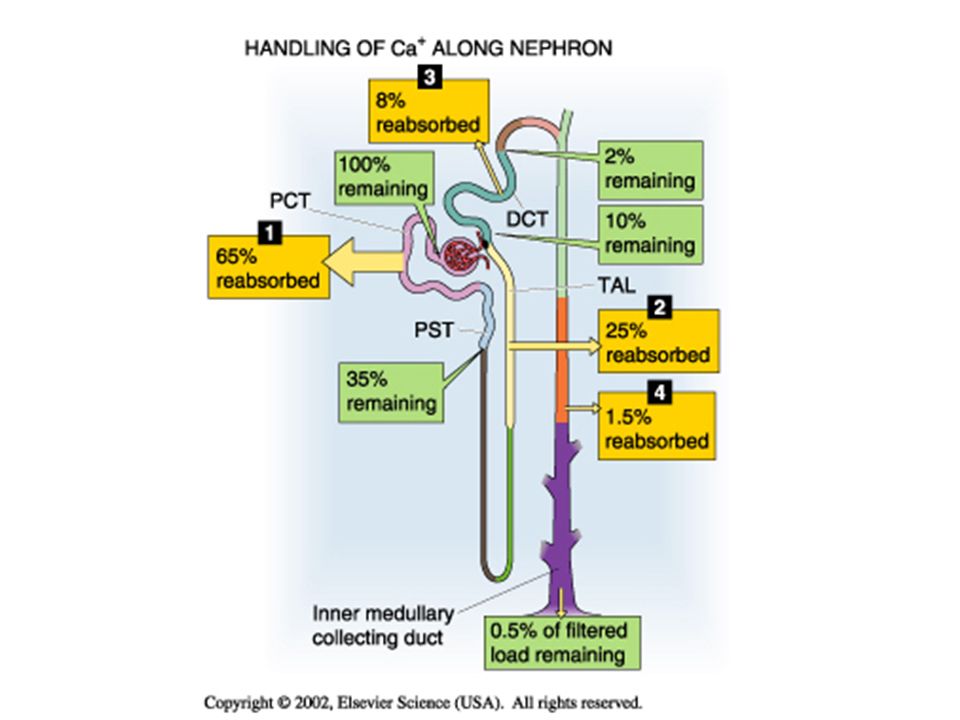
The renal tubule has the ability to vary its reabsorption of filtered calcium to adjust for changes in intake or increased calcium requirements. Under normal conditions, <1% of filtered calcium is excreted in the urine. Calcium is reabsorbed throughout the nephron; Most of the regulation of reabsorption occurs in the distal nephron. In the thick ascending limb of the loop of Henle (TAL), calcium reabsorption occurs through a primarily paracellular pathway, but active, transcellular reabsorption may occur as well during stimulation with parathyroid hormone (PTH).
, calcium reabsorption occurs through a primarily paracellular pathway, but active, transcellular reabsorption may occur as well during stimulation with parathyroid hormone (PTH)..")
66
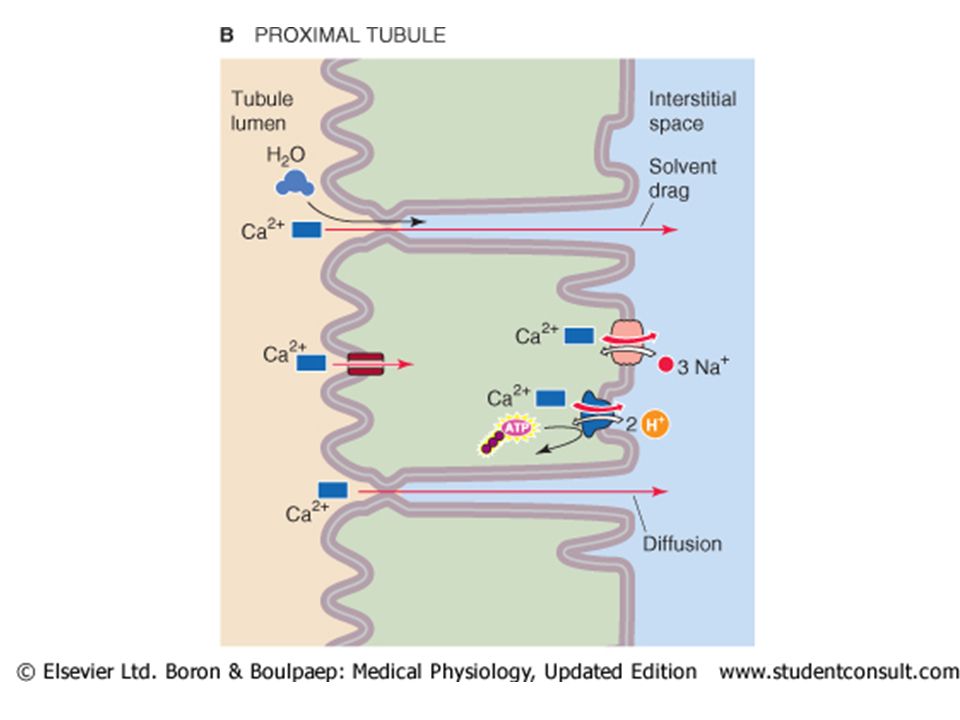
Calcium reabsorption in the PROXIMAL TUBULE. The proximal tubule reabsorbs approximately 65% of the filtered Ca 2+, a process that is not subject to hormonal control. A small part of the Ca 2+ reabsorbed by the proximal tubule (20% of the 65%) moves via a transcellular route. Most proximal tubule Ca 2+ reabsorption (80% of the 65%) occurs via the paracellular route.
 moves via a transcellular route. Most proximal tubule Ca 2+ reabsorption (80% of the 65%) occurs via the paracellular route..")
68
Calcium Transport in the THICK ASCENDING LIMB. The thick ascending limb (TAL) reabsorbs approximately 25% of the filtered Ca 2+. Under normal conditions, about half of Ca 2+ reabsorption in the TAL occurs passively via a paracellular route, driven by the lumen-positive voltage. Thus, it is not surprising that hormones such as AVP, which make the transepithelial voltage more positive, indirectly increase Ca 2+ reabsorption. The other half of Ca 2+ reabsorption by the TAL occurs via the transcellular pathway, which is stimulated by PTH.
 reabsorbs approximately 25% of the filtered Ca 2+. Under normal conditions, about half of Ca 2+ reabsorption in the TAL occurs passively via a paracellular route, driven by the lumen-positive voltage. Thus, it is not surprising that hormones such as AVP, which make the transepithelial voltage more positive, indirectly increase Ca 2+ reabsorption. The other half of Ca 2+ reabsorption by the TAL occurs via the transcellular pathway, which is stimulated by PTH..")
69
CaSR on basal-lateral surface may inhibit Ca transport and NaKCl co- transport Calcium enters the apical side passively, and most is then extruded via active transport out of the cell into the basal lateral interstitium.

70
Calcium in the DISTAL CONVOLUTED TUBULE. This segment reabsorbs approximately 8% of the filtered Ca 2+ load. Despite the relatively small amount of Ca 2+ delivered, the distal convoluted tubule (DCT) is a major regulatory site for Ca 2+ excretion. In contrast to the proximal tubule and TAL, the DCT reabsorbs Ca 2+ predominantly via an active, transcellular route.
 is a major regulatory site for Ca 2+ excretion. In contrast to the proximal tubule and TAL, the DCT reabsorbs Ca 2+ predominantly via an active, transcellular route..")
71
PARATHYROID HORMONE The most important regulator of renal Ca2+ reabsorption is PTH, which stimulates Ca2+ reabsorption in the thick ascending limb, the distal convoluted tubule, and the connecting tubule. (PTH does NOT have a proximal action) PTH appears to increase the open probability of apical Ca2+ channels. Such an increase in Ca2+ permeability would increase intracellular [Ca2+]i, which in turn would stimulate basolateral Ca2+ extrusion mechanisms, increase Ca2+ reabsorption, and raise plasma [Ca2+ ].
 PTH appears to increase the open probability of apical Ca2+ channels. Such an increase in Ca2+ permeability would increase intracellular [Ca2+]i, which in turn would stimulate basolateral Ca2+ extrusion mechanisms, increase Ca2+ reabsorption, and raise plasma [Ca2+ ]..")
72
Calcium reabsorption in the Distal Convoluted Tubule (DCT) Because only a small percentage of filtered calcium is excreted in the urine (0.5%), and the DCT is the last major sight for reabsorption of calcium, relatively small changes in the fraction reabsorbed in the DCT result in large changes in the total amount of calcium lost in the urine. Reabsorption in the DCT is regulated by PTH. When serum calcium concentration decreases, PTH release from the parathyroid glands increases. This effect is mediated by the calcium-sensing receptor (CaSR) in the parathyroid gland. PTH acts on the DCT to increase calcium reabsorption.
 in the parathyroid gland. PTH acts on the DCT to increase calcium reabsorption..")
73
K +, Potassium The distribution of potassium is very different from sodium, as it is the most abundant intracellular cation, as 98% of the bodies K + lies within cells. The circulating K + is tightly regulated at a low 3.5-5.0 mM (compared to 145 mM for Na + ). K + within cells is essential for maintaining cell volume, regulating intracellular pH, controlling enzyme function, DNA, protein synthesis and cell growth. The ratio of intracellular to low extracellular K + is critical in maintaining the electrical potential across cell membrane (in both excitable and non-excitable cells).
. K + within cells is essential for maintaining cell volume, regulating intracellular pH, controlling enzyme function, DNA, protein synthesis and cell growth. The ratio of intracellular to low extracellular K + is critical in maintaining the electrical potential across cell membrane (in both excitable and non-excitable cells)..")
74
Potassium homeostasis: The body must remain in “K + balance,” such that intake equals output. (about 80-120 mM K + /day; in and out). Typically the kidney filters 800 mM/day, so the excretory load is about 10-15% of that filtered. K + handling and regulation by the kidney includes the ability both to reabsorb and to secrete.
. Typically the kidney filters 800 mM/day, so the excretory load is about 10-15% of that filtered. K + handling and regulation by the kidney includes the ability both to reabsorb and to secrete..")
75
90% of filtered K is reabsorbed by the end of the loop of Henle. Regulation of K+ balance depends on the 5 distal nephron segments

76
Most (80%) of K + reabsorption takes place in the Proximal tubule Proximal reabsorption of 80% of the filtered load takes place by passive paracellular movement via solvent drag and simple diffusion, driven by the gradients established by active sodium reabsorption. Potassium reabsorption in the proximal (because of its passive paracellular nature) is highly dependent on fluid movement out of the lumen, following sodium. The basalateral Na-K pump does NOT directly affect K+ rabsorption since the movement is mostly paracellular and not dependent on the intracellular gradient (remember that the [K] i is high and works against transcellular K+ movement!)
 is highly dependent on fluid movement out of the lumen, following sodium. The basalateral Na-K pump does NOT directly affect K+ rabsorption since the movement is mostly paracellular and not dependent on the intracellular gradient (remember that the [K] i is high and works against transcellular K+ movement!).")
77
K+ reabsorption in the thick ascending limb accounts for an unregulated additional 10% of the filtered load. The thin limbs of the loop of Henle secrete K+ (passively) due to the high K concentration in the medullary interstitium. The TAL reabsorbs K + by both paracellular and transcellular pathways, Paracellular passive reabsorption is driven by both high K permeability and the lumen positive voltage. Transcellular reabsorption is driven by the NKCC2 co- transporter moving sodium and K into the cell via the apical membrane, driven by the active movement of Na (due to the basalateral Na-K pump) across the cell.
 due to the high K concentration in the medullary interstitium. The TAL reabsorbs K + by both paracellular and transcellular pathways, Paracellular passive reabsorption is driven by both high K permeability and the lumen positive voltage. Transcellular reabsorption is driven by the NKCC2 co- transporter moving sodium and K into the cell via the apical membrane, driven by the active movement of Na (due to the basalateral Na-K pump) across the cell..")
78
Paracellular = 50% Transcellular K+ 50%

79
In the collecting ducts, the two cell types have different functions: The principal cells secrete K+ Secretion requires a basalateral Na-K exchanger to let K enter the cell against it gradient, and K channels on the apical side to allow K+ to flow into the lumen, down its chemical gradient. The intercalated cells reabsorb K+ Reabsorption requires an apical K+ pump to push K into the cell, and K channels on the basalateral membrane to allow K to leave the cell down its chemical gradient.

80
K+ SecretionK+ Reabsorption

Similar presentations










 and, >")



, the thin descending and.>")




. Other excretory.>")