Download presentation
Presentation is loading. Please wait.

1
Biochemistry I Chapter 11 Enzymatic Catalysis Dr. Loren Williams
Revised 4/30/2014 Biochemistry I Dr. Loren Williams Chapter 11 Enzymatic Catalysis

2
Chapter 11 Opener

3
ENZYMES high reaction rates,
controllable reaction rates (cooperativity, allosterism) high specificity, mild conditions. Table 11-1
 high specificity, mild conditions. Table")
4
Table 11-2

5
Enzymes bind to specific substrates, via non-covalent interactions
complementarity of surfaces, and interactions, sometimes ‘lock and key’ sometimes ‘induced fit’ with stereospecificity generally but not always with chemical (geometric) specificity peptidases versus esterases Figure 11-1
 specificity. peptidases versus esterases. Figure")
6
Prochirality: A tetrahedral carbon that can be converted to a chiral carbon by changing any one of its attached groups is ‘prochiral’. All the substitutuents of a prochiral carbon are distinguishable in a chiral environment (in the vicinity of any biological macromolecule). The brown and the blue functional groups behave identically in an achiral environment. In a chiral environment the brown and the blue functional groups are different and can be distinguished. These two can be distinguished by an enzyme: they occupy different regions in three-dimensional space. Prochirality is an important concept in biological chemistry and was discovered by biochemists.
. The brown and the blue functional groups behave identically in an achiral environment. In a chiral environment the brown and the blue functional groups are different and can be distinguished. These two can be distinguished by an enzyme: they occupy different regions in three-dimensional space. Prochirality is an important concept in biological chemistry and was discovered by biochemists.")
7
A prochiral molecule in chiral environment - is chiral.
potentially chiral nearly asymmetric Figure 11-2

8
This molecule is prochiral, the two faces are different, only when it is in a chiral environment. Otherwise, they are the same.

9
Some enzymes are not too specific (this is unusual).
Page 326

10
Figure 11-3

12
NAD or NADP NADH or NADPH

13
Schematic structures of distinct identifying domains in Nox isoform catalytic subunits.
Schematic structures of distinct identifying domains in Nox isoform catalytic subunits. A: Nox2 and Nox2-like isoforms (Nox1, 3, and 4) present a basic structure of 6 transmembrane domains containing two heme groups and COOH-terminal FAD and NADPH domains. B: in addition to those domains, Nox5 possesses Ca2+-binding EF motifs in the NH2-terminal region. C: dual oxidasess, in addition to Ca2+-binding EF motifs, possess an NH2-terminal peroxidase-like domain. NADPH Oxidases: Schematic structures of distinct identifying domains in Nox isoform catalytic subunits. A: Nox2 and Nox2-like isoforms (Nox1, 3, and 4) present a basic structure of 6 transmembrane domains containing two heme groups and COOH-terminal FAD and NADPH domains. B: in addition to those domains, Nox5 possesses Ca2+-binding EF motifs in the NH2-terminal region. C: dual oxidasess, in addition to Ca2+-binding EF motifs, possess an NH2-terminal peroxidase-like domain. Frazziano G et al. Am J Physiol Heart Circ Physiol 2012;302:H2166-H2177
 present a basic structure of 6 transmembrane domains containing two heme groups and COOH-terminal FAD and NADPH domains. B: in addition to those domains, Nox5 possesses Ca2+-binding EF motifs in the NH2-terminal region. C: dual oxidasess, in addition to Ca2+-binding EF motifs, possess an NH2-terminal peroxidase-like domain. NADPH Oxidases: Schematic structures of distinct identifying domains in Nox isoform catalytic subunits. A: Nox2 and Nox2-like isoforms (Nox1, 3, and 4) present a basic structure of 6 transmembrane domains containing two heme groups and COOH-terminal FAD and NADPH domains. B: in addition to those domains, Nox5 possesses Ca2+-binding EF motifs in the NH2-terminal region. C: dual oxidasess, in addition to Ca2+-binding EF motifs, possess an NH2-terminal peroxidase-like domain. Frazziano G et al. Am J Physiol Heart Circ Physiol 2012;302:H2166-H2177.")
14
reduced oxidized oxidized reduced Page 327

15
Figure 11-5a

18
Figure 11-5b

19
What does an enzyme do to this graph?

21
Figure 11-6

22
Figure 11-7

23
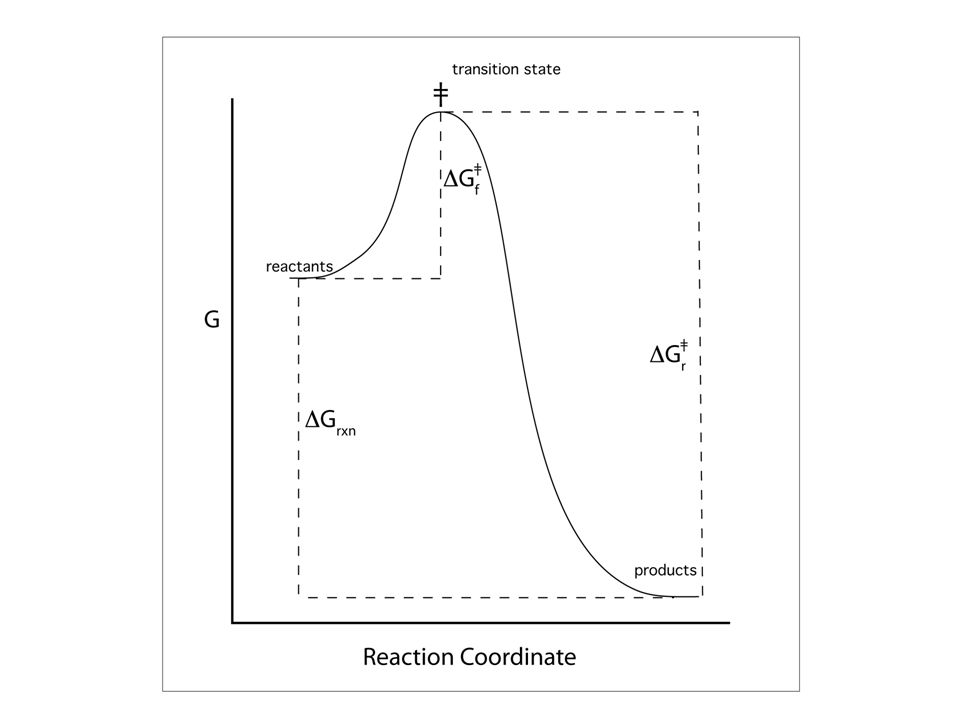
DG°rxn= DH°rxn-TDS°rxn K = kr/ff ; DG°rxn = DG°‡r- DG°‡f
Relationships between rate constants equilibrium constants activation free energy free energy of reaction DG°rxn=-RTlnK DG°rxn= DH°rxn-TDS°rxn K = kr/ff ; DG°rxn = DG°‡r- DG°‡f

24
The Transition State The highest energy species (most unstable species) along the reaction coordinate.
 along the reaction coordinate.")
25
Page 330

26
uncatalyzed acid catalyzed base catalyzed Figure 11-8

27
Figure 11-8a

28
Figure 11-8b

29
Figure 11-8c

30
Effect of pH on catalytic activity.
This profile suggests a histidine is involved in the mechanism. Box 11-1

31
Elements of Catalysis acids & bases covalent intermediates metal ions
proximity and orientation (VVP calls this section ‘mechanisms of catalysis, that seems a bit illogical. These are components or elements of catalysis. Catalytic mechanisms incorporate one or more of these elements.)
")
32
RNAse H an acid and a base Figure 11-9

33
RNAse H E + S ⇌ ES ⇌ EI ⇌ EP ⇌ E + P Figure 11-10

34
Figure part 1

35
Figure part 2

36
Covalent Intermediates
enzyme nucleophile Page 334

37
Covalent Intermediates
uncatalyzed reaction (nucleophile on enzyme) catalyzed reaction Figure 11-11
 catalyzed reaction. Figure")
38
Covalent Intermediates
Figure 11-12

39
Metal ions in catalysis:
Carbonic Anhydrase The carbonic anhydrase, a metalloenzyme (contains a Zn2+) catalyzes the conversion of bicarbonate and a proton to carbon dioxide and water also catalyzes the the reverse: conversion of carbon dioxide and water to bicarbonate and a proton is a very efficient enzyme causes carbonated drinks to rapidly degas in your mouth the Zn2+ is square planer is coordinated by three His plus one water molecule the water molecule is polarized by the Zn and is highly acidic Figure 11-13a
 catalyzes the conversion of bicarbonate and a proton to carbon dioxide and water. also catalyzes the the reverse: conversion of carbon dioxide and water to bicarbonate and a proton. is a very efficient enzyme. causes carbonated drinks to rapidly degas in your mouth the Zn2+ is square planer is coordinated by three His. plus one water molecule. the water molecule is polarized by the Zn and is highly acidic. Figure 11-13a.")
40
Figure 11-13b

41
Proximity and Orientation: Entropy Trapping
(i) Proximity increases reaction rates Slower Faster Page 336
 Proximity increases reaction rates. Slower. Faster. Page 336.")
42
(ii) Orientation increases reaction rates:
(i.e., correct + fixed orientation increases reaction rates.) For example an SN2 reaction reaction is fastest if the attacking nucleophile approaches the electrophilic C along the line of the leaving group bond, from the opposite side of the C. Figure 11-14
 For example an SN2 reaction reaction is fastest if the attacking nucleophile approaches the electrophilic C along the line of the leaving group bond, from the opposite side of the C. Figure")
43
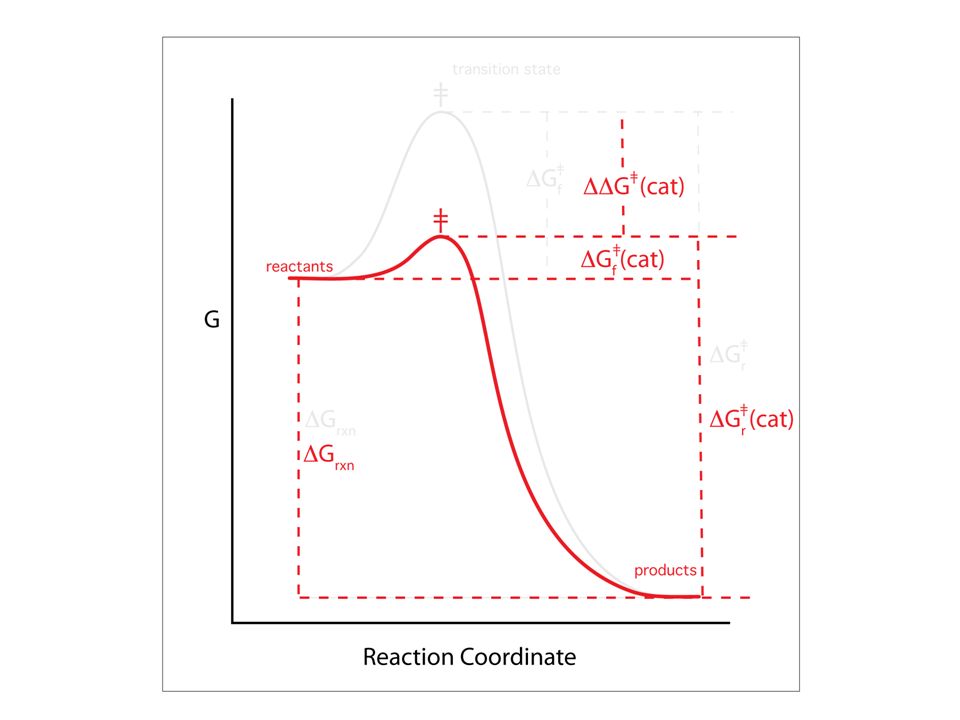
Preferential Binding to Transition States
The transition state can be stabilized by mechanically distorting the substrate to a non-ground state conformation that is close to the transition state conformation. This example shows how a non-enzymatic transition state can be stabilized (Big R-group lowers DG‡) Page 338
 Page 338.")
44
Binding to Transition States
I don’t’ like this graph, which is intended to relate to preferential binding of the enzyme to the transition state. In fact, all the elements of catalysis discussed in this section (acid base, metals, proximity and orientation) will lower DG‡ and will contribute to a graph shape like this. The small double headed arrow is the binding free energy of the substrate to the enzyme. I think that to be specifically related to binding to the transition state, this graph should have H (enthalpy) on the vertical axis and should explain that the binding energy can be converted to mechanical distortion of the substrate toward the transition state. E + S ⇌ ES ⇌ EP ⇌ E + P Figure 11-15
 will lower DG‡ and will contribute to a graph shape like this. The small double headed arrow is the binding free energy of the substrate to the enzyme. I think that to be specifically related to binding to the transition state, this graph should have H (enthalpy) on the vertical axis and should explain that the binding energy can be converted to mechanical distortion of the substrate toward the transition state. E + S ⇌ ES ⇌ EP ⇌ E + P. Figure")
45
Transistion State Analogs
‡

46
Transistion State Analogs
‡

47
Lysozyme Lysozyme breaks up peptidoglycans, which form the cell walls of bacteria. Lysozyme hydrolyzes the glycosidic bond that connects the 1-oxygen of N-acetylmuramic acid with the 4- carbon of N-acetylglucosamine. Lysozyme conferes protection from bacterial infection. For example the membrane covering the eye is protected by secreted lysozyme and defensin. The 3D structure of chicken egg-white lysozyme was determined to 2 Å resolution by X-ray diffraction (Phillips, 1965). Lysozyme was the second protein structure determined (after myoglobin, Perutz & Kendrew) and was the first enzyme whose structure was solved. Lysozyme was therefore the first enzyme for which the detailed mechanism was known.
. Lysozyme was the second protein structure determined (after myoglobin, Perutz & Kendrew) and was the first enzyme whose structure was solved. Lysozyme was therefore the first enzyme for which the detailed mechanism was known.")
48
The following slides are the peptidoglycan section in chapter 8 of VVP (page ) in the 4th edition. You must know this material for the next exam

49
Peptidogycan are stained by Gram staining are not stained by Gram staining

50
Plant cell walls are made from cellulose
Plant cell walls are made from cellulose. Bacterial cell walls are made from peptidoglycan. Gram-positive cell walls have a high amount of peptidoglycan in their cell walls and lack outer membranes, unlike in Gram-negative bacteria. Figure 8-16a

51
Gram negative bacteria contain an outer membrane composed of lipopolysaccharide which contains porins (hole-making proteins). The space between the peptidoglycan and the outer membrane is called the periplasmic space.

53
N-Acetyl-D-Glucosamine GlcNAc NAM N-acetylmuramic acid
Peptidoglycan - carbohydrates + peptides carbohydrates: alternating N-acetylglucosamine (NAM) and N-acetylmuramic acid (NAG) with β-(1,4) linkages. Peptides - oligopeptides (3-5 AA long) linked to N-acetylmuramic acids, and cross-linked to other oligopeptides (pentaglycine) The peptide chain cross-links to the peptide chain of another strand forming the 3D mesh-like layer. Peptidoglycan serves a structural role in the bacterial cell wall, giving structural strength, as well as counteracting the osmotic pressure of the cytoplasm. NAG N-Acetyl-D-Glucosamine GlcNAc NAM N-acetylmuramic acid
 and N-acetylmuramic acid (NAG) with β-(1,4) linkages. Peptides - oligopeptides (3-5 AA long) linked to N-acetylmuramic acids, and cross-linked to other oligopeptides (pentaglycine) The peptide chain cross-links to the peptide chain of another strand forming the 3D mesh-like layer. Peptidoglycan serves a structural role in the bacterial cell wall, giving structural strength, as well as counteracting the osmotic pressure of the cytoplasm. NAG. N-Acetyl-D-Glucosamine GlcNAc. NAM. N-acetylmuramic acid.")
54
Figure 8-17a

56
Penicillin was discovered by Alexander Fleming in 1928
Penicillin was discovered by Alexander Fleming in He showed the fungus Penicillium notatum can exude a substance with antibiotic properties, which he dubbed penicillin. In one of the best examples of the power of basic research, Fleming's 'serendipitous' discovery (from a discarded, contaminated Petri dish) has saved 100’s of millions of lives. Penicillin prevents formation of peptidoglycan cross-links by inhibiting the the enzyme (DD-transpeptidase).
 has saved 100’s of millions of lives. Penicillin prevents formation of peptidoglycan cross-links by inhibiting the the enzyme (DD-transpeptidase).")
58
Back to lysozyme

59
E D N-acetylmuramic acid N-acetylglucosamine Figure 11-16

60
Figure 11-17

61
Figure 11-19

62
Identifying the cleaved bond and the products: retention of configuration.
18-O at C1 or C4? Page 343

63
oxonium ion-carbocation: trigonal planer intermediate
non-enzymatic cleavage Figure 11-20

64
Figure 11-21

65
(ignore the electron flow arrows in this slide, that comes later)
The enzyme binds to the substrate and some of the binding energy is used to distort the D-ring to the half chair. Therefore the energy of binding is used to stabilize the transition state. (ignore the electron flow arrows in this slide, that comes later) D Figure part 1
 D. Figure part 1.")
66
Proton transfer to the bridging O1 atom. Now the electrons flow.
cleavage trigonal planer intermediate D D Figure part 2

67
nucleophilic attack by Asp 52
D D D tetrahedral intermediate, covalent E-S intermediate Figure part 3

68
Ignore the electron flow on this panel
Water replaces the E ring in the active site. Figure part 4

69
4) And ASP35 – substrate bond cleaves
5) Then, product release. 4) And ASP35 – substrate bond cleaves 3) And, base-catalysis (Glu35 accepts a proton from the water). 2) Then, nucleophilic attack by water at the C1 of the D ring 1) After water replaces the E ring in the active site (previous slide_. Figure part 5
 Then, product release. 4) And ASP35 – substrate bond cleaves. 3) And, base-catalysis (Glu35 accepts a proton from the water). 2) Then, nucleophilic attack. by water at the C1. of the D ring. 1) After water replaces the. E ring in the active site (previous slide_. Figure part 5.")
70
Figure 11-22

71
(no acid/base catalysis) NAG2FGLcF F: magenta atom
E35Q (no acid/base catalysis) NAG2FGLcF F: magenta atom (stabilizes the carbocation) Figure 11-23
 NAG2FGLcF. F: magenta atom. (stabilizes the carbocation) Figure")
72
Serine proteases cut peptide bonds in proteins via nucleophilic attack by a serine.
Are grouped into clans that share structural similarities (homology) chymotrypsin-like, subtilisin-like, alpha/beta hydrolase, signal peptidase clans. Ancestral serine proteases in mammels were digestive enzymes that evolved by gene duplication and now function in blood clotting, the immune system, and inflammation. Serine proteases are paired with serine protease inhibitors.
 chymotrypsin-like, subtilisin-like, alpha/beta hydrolase, signal peptidase clans. Ancestral serine proteases in mammels were digestive enzymes that evolved by gene duplication and now function in blood clotting, the immune system, and inflammation. Serine proteases are paired with serine protease inhibitors.")
73
not covered Page 348

74
not covered Figure 11-24

75
The nucleophilic serine can be identified by chemical labeling.
not covered The nucleophilic serine can be identified by chemical labeling. Page 348

76
Acetylcholine is aneurotransmitter.
Accumulation of acetylcholine causes continuous stimulation of the muscles, glands, and central nervous system. DIPF, Sarin and VX nerve gas are acetylcholine esterase inhibitors. Box 11-3a

77
Box 11-3c

78
acetylcholine esterase inhibitors
Box 11-3b

79
Figure 11-25

80
Figure 11-26

81
Specificity pockets Figure 11-27

82
Not covered Figure 11-28

84
X X Figure 11-29

85
X Figure part 1

86
X Figure part 2

87
Figure part 3

88
X Figure part 4

89
X Figure part 5

90
Figure 11-30a

91
Figure 11-30b

92
BPTI: bovine pancreatic trypsin inhibitor
BPTI is a member of the protein family of Kunitz-type serine protease inhibitors. Its primary function seems to be inhibition of trypsin in the pancreas. Small amounts of trypsinogen are cleaved during storage in the pancreas (not good). Wikipedia: BPTI is one of the most thoroughly studied proteins by structural biology methods, experimental and computational dynamics, mutagenesis, and folding experiments. It was one of the earliest protein crystal structures solved, in 1970 in the laboratory of Robert Huber, and was the first protein to have its structure determined by NMR spectroscopy. It was first macromolecule of scientific interest to be simulated using molecular dynamics. Figure 11-31a
. Wikipedia: BPTI is one of the most thoroughly studied proteins by structural biology methods, experimental and computational dynamics, mutagenesis, and folding experiments. It was one of the earliest protein crystal structures solved, in 1970 in the laboratory of Robert Huber, and was the first protein to have its structure determined by NMR spectroscopy. It was first macromolecule of scientific interest to be simulated using molecular dynamics. Figure 11-31a.")
93
Trypsin BPTI Figure 11-31b

94
elastase, ph 9.0, quick freeze:
Figure 11-32

95
Figure 11-32a

96
Figure 11-32b

97
Trypsin is dangerous and is synthesized as an inactive form called a zymogen. The zymogen of trypsin is called trypsinogen, which is found in pancreatic juice, along with amylase, lipase, chymotrypsinogen, nucleases, etc. Trypsinogen is activated (converted to trypsin) by cleavage after amino acid 15 by the protease enteropeptidase. Enteropeptidase is found in the intestinal mucosa. In addition, Trypsin can cleave trypsinogen to form trypsin. Figure 11-33

98
Coagulation Cascade Box 11-4a

99
Factor XIa is a serine protease that is synthesized as a zymogen called factor XI.
Factor XIa activates Factor IX by proteolysis to give Factor IXa. Factor IXa is a serine protease (synthesized as zymogen Factor IX). Factor IXa activates Factor X by proteolysis to give Factor Xa Factor Xa is a serine protease (synthesized as zymogen Factor X). The activity of factor Xa is enhanced by Factor V. Factor Xa cleaves prothrombin to form thrombin. Thrombin is a serine protease (synthesized as zymogen prothrombin). image from promega
. Factor IXa activates Factor X by proteolysis to give Factor Xa. Factor Xa is a serine protease (synthesized as zymogen Factor X). The activity of factor Xa is enhanced by Factor V. Factor Xa cleaves prothrombin to form thrombin. Thrombin is a serine protease (synthesized as zymogen prothrombin). image from promega.")
100
Box 11-4b

101
Hemophilia A is clotting factor VIII deficiency (1 in 5,000–10,000 male births.)
Hemophilia B is factor IX deficiency (1 in about 20,000–34,000 male births). Box 11-4c
. Box 11-4c.")
102
Protease Classes Serine proteases Threonine proteases
Cysteine proteases Aspartate proteases Glutamic acid proteases Metalloproteases Enzymes that catalyse the hydrolytic cleavage of peptide bonds are called proteases. Proteases fall into four main mechanistic classes: serine, cysteine, aspartyl and metalloproteases. In the active sites of serine and cysteine proteases, the eponymous residue is usually paired with a proton-withdrawing group to promote nucleophilic attack on the peptide bond. Aspartyl proteases and metalloproteases activate a water molecule to serve as the nucleophile, rather than using a functional group of the enzyme itself. However, the overall process of peptide bond scission is essentially the same for all protease classes. Soluble serine proteases (a); cysteine proteases (b); aspartyl proteases (c); and metalloproteases (d). © 2009 Nature Publishing Group
; cysteine proteases (b); aspartyl proteases (c); and metalloproteases (d). © 2009 Nature Publishing Group.")
Similar presentations