Download presentation
Presentation is loading. Please wait.

1
ENZYMES

2
History of Enzymes -1700s and early 1800s, the digestion of meat by stomach secretions and the conversion of starch to sugars by plant extracts and saliva were known. --mechanism by which this occurred had not been identified.

3
History of Enzymes -19th century, when studying the fermentation of sugar to alcohol by yeast, Louis Pasteur came to the conclusion that it was catalyzed by a vital force contained within the yeast cells called "ferments", which were thought to function only within living organisms. --He wrote that "alcoholic fermentation is an act correlated with the life and organization of the yeast cells, not with the death or putrefaction of the cells. Yeast

4
History of Enzymes 1878: German physiologist Wilhelm Kühne
first used the term enzyme, which literally means “in yeast” 1897: Eduard Buchner began to study the ability of yeast extracts that lacked any living yeast cells to ferment sugar. He also found that the sugar was fermented even when there were no living yeast cells in the mixture. He named the enzyme that brought about the fermentation of sucrose "zymase". In 1907 he received the Nobel Prize in Chemistry “for his biochemical research and his discovery of cell-free fermentation".

5
Enzymes Enzymes are biomolecules that catalyze, increase the rates of chemical reactions without being altered during the reaction. Almost all enzymes are proteins; Enzymes are essential to life. In enzymatic reactions, the molecules at the beginning of the process are called substrates, and the enzyme converts them into different molecules, the products.

6
Enzymes Almost all processes in a biological cell need enzymes in order to occur at significant rates. Since enzymes are extremely selective for their substrates and speed up only a few reactions from among many possibilities, the set of enzymes made in a cell determines which metabolic pathways occur in that cell.

7
Enzymes Enzyme activity can be affected by other molecules.
Inhibitors are molecules that decrease enzyme activity. Inducers are molecules that increase activity. Many drugs and poisons are enzyme inhibitors. Activity is also affected by temperature, chemical environment (e.g. pH),
,")
8
Enzyme Inhibitor & Inducer
Enzyme Inducer Cimetidine Rifampicin Ketoconazole Carbamazepine Fluconazole Phenobarbital Miconazole Phenytoin Macrolides(except Azithromycin) Griseofulvin Fluoroquinolones(except Levofloxacin) Smoking Chronic alcoholism
 Griseofulvin. Fluoroquinolones(except Levofloxacin) Smoking. Chronic alcoholism.")
9
ENZYMES are biological catalyst
ENZYME CHARACTERISTICS The basic function of an enzyme is to increase the rate of a reaction Most enzymes act specifically with only one reactant (called a substrate) to produce products The most remarkable characteristic is that enzymes are regulated from a state of low activity to high activity and vice versa
 to produce products. The most remarkable characteristic is that enzymes are regulated from a state of low activity to high activity and vice versa.")
10
Enzymes Lower a Reaction’s Activation Energy
Energy and Enzymes 4/22/2017 Enzymes Lower a Reaction’s Activation Energy G. Podgorski, Biol. 1010

11
Energy and Enzymes 4/22/2017 Enzyme Action G. Podgorski, Biol. 1010

12
Three-dimensional structure of an ENZYME

13
Three-dimensional structure of an ENZYME
Enzymes are proteins They have a globular shape A complex 3-D structure Human pancreatic amylase

14
Enzyme Structure Most enzymes are proteins
Enzymes may require a non-peptide component as a cofactor. The peptide component is called the apoenzyme, the cofactor is called as the coenzyme and the combined functional unit is the holoenzyme Cofactors that are tightly bound to the polypeptide are called prosthetic groups. Such proteins are called as complex or conjugated proteins. Proteins without prosthetic groups are simple proteins

15
The Active Site One part of an enzyme, the active site, is particularly important The shape and the chemical environment inside the active site permits a chemical reaction to proceed more easily

16
APOENZYME PROENZYME OR ZYMOGEN The inactive form of the apoenzyme
May be inactive in its original synthesized structure PROENZYME OR ZYMOGEN The inactive form of the apoenzyme May contain several extra amino acids in the protein which are removed, and allows the final specific tertiary structure to be formed before it is activated as an apoenzyme APOENZYME

17
The Substrate The substrate of an enzyme are the reactants that are activated by the enzyme; Enzymes are specific to their substrates; The specificity is determined by the active site.

18
COFACTOR An additional non-protein molecule that is needed by some enzymes to help the reaction Nitrogenase enzyme with Fe, Mo and ADP cofactors

19
COFACTOR A non-protein substance which may be organic and called coenzyme Common coenzymes are vitamins and metal ions

20
COFACTOR Another type of cofactor is an inorganic metal ion called a metal ion activator Are inorganic and may be bonded through coordinate covalent bonds Metal ions as Zn+2, Mg+2, Mn+2, Fe+2, Cu+2, K+, and Na+1 are used in enzymes as cofactors

22
Vitamins as Coenzymes Vitamin Coenzyme Function Niacin
nicotinamide adenine dinucleotide (NAD+) oxidation or hydrogen transfer Riboflavin flavin adenine dinucleotide (FAD) Pantothenic acid coenzyme A (CoA) Acetyl group carrier Vitamin B12 coenzyme B-12 Methyl group transfer Thiamine (B1) thiaminpyrophosphate (TPP) Aldehyde group transfer
 oxidation or hydrogen transfer. Riboflavin. flavin adenine dinucleotide (FAD) Pantothenic acid. coenzyme A (CoA) Acetyl group carrier. Vitamin B12. coenzyme B-12. Methyl group transfer. Thiamine (B1) thiaminpyrophosphate (TPP) Aldehyde group transfer.")
23
PROSTHETIC GROUPS Are tightly incorporated into protein structure by covalent or noncovalent forces Examples include derivatives of B vitamins such as pyridoxal phosphate, flavin mononucleotide (FMN), flavin adenine dinucleotide (FAD), thiamin pyrophosphate, biotin and METAL IONS of Co, Cu, Mg, Mn, and Zn. METALLOENZYMES – enzymes that contain tightly bound metal ions
, flavin adenine dinucleotide (FAD), thiamin pyrophosphate, biotin and METAL IONS of Co, Cu, Mg, Mn, and Zn. METALLOENZYMES – enzymes that contain tightly bound metal ions.")
24
PROSTHETIC GROUPS

25
NOMENCLATURE The commonly used names for most enzymes describe the type of reaction catalyzed, followed by the suffix –ase. Dehydrogenases – remove hydrogen atoms Proteases – hydrolyze proteins Isomerases – catalyze rearrangement in configuration

26
NOMENCLATURE Modifiers may precede the name to indicate;
(a) the substrate (xanthine oxidase) (b) the source of the enzyme (pancreatic ribonuclease) (c) its regulation (hormone-sensitive lipase) (d) a feature of its mechanism of action (cysteine protease)
 the substrate (xanthine oxidase) (b) the source of the enzyme (pancreatic ribonuclease) (c) its regulation (hormone-sensitive lipase) (d) a feature of its mechanism of action (cysteine protease)")
27
NOMENCLATURE Alphanumeric designators may be added to identify multiple forms of an enzyme ( eg., RNA polymerase III; protein kinase C ) Some enzymes retain their original trivial names, which give no hint of the associated enzymatic reaction Examples are pepsin, trypsin, and chymotrypsin which catalyzes the hydrolysis of proteins

29
Classification of Enzymes
Based on catalyzed reactions, the nomenclature committee of the International Union of Biochemistry and Molecular Biology (IUBMB) recommended the following classification: OXIDOREDUCTASES Catalyze a variety of oxidation-reduction reactions Common names include dehydrogenase, oxidase, reductase and catalase
 recommended the following classification: OXIDOREDUCTASES. Catalyze a variety of oxidation-reduction reactions. Common names include dehydrogenase, oxidase, reductase and catalase.")
30
Classification of Enzymes
2. TRANSFERASES Catalyze transfers of groups (acetyl, methyl, phosphate, etc.). The first three subclasses play major roles in the regulation of cellular processes. The polymerase is essential for the synthesis of DNA and RNA.
. The first three subclasses play major roles in the regulation of cellular processes. The polymerase is essential for the synthesis of DNA and RNA.")
31
Three major regulatory chemical reactions
Three major regulatory chemical reactions. (a) Acetylation - addition of an acetyl group to lysine's R group by acetyltransferase. (b) Methylation - addition of a methyl group to DNA's base (e.g. cytosine) by methylase. (c) Phosphorylation - addition of a phosphate group to the R group of tyrosine, serine or threonine (only tyrosine is shown here) by protein kinase.
 Acetylation - addition of an acetyl group to lysine s R group by acetyltransferase. (b) Methylation - addition of a methyl group to DNA s base (e.g. cytosine) by methylase. (c) Phosphorylation - addition of a phosphate group to the R group of tyrosine, serine or threonine (only tyrosine is shown here) by protein kinase.")
32
Classification of Enzymes
3. HYDROLASES Catalyze hydrolysis reactions where a molecule is split into two or more smaller molecules by the addition of water PROTEASES split protein molecules HIV protease is essential for HIV replication Caspase plays a major role in apoptosis NUCLEASES split nucleic acids (DNA and RNA) Based on the substrate type, they are divided into RNase and DNase. RNase catalyzes the hydrolysis of RNA DNase acts on DNA
 Based on the substrate type, they are divided into RNase and DNase. RNase catalyzes the hydrolysis of RNA. DNase acts on DNA")
33
Classification of Enzymes
Nucleases cont… They may also be divided into exonuclease and endonuclease. The exonuclease progressively splits off single nucleotides from one end of DNA or RNA. The endonuclease splits DNA or RNA at internal sites. PHOSPHATASE catalyzes dephosphorylation (removal of phosphate groups). Example: calcineurin (also known as protein phosphatase 3) The immunosuppressive drugs Tacrolimus, Sirolimus, Everolimus and Cyclosporin A are the calcineurin inhibitors
. Example: calcineurin (also known as protein phosphatase 3) The immunosuppressive drugs Tacrolimus, Sirolimus, Everolimus and Cyclosporin A are the calcineurin inhibitors.")
34
Classification of Enzymes
4. LYASES Catalyze the cleavage of C-C, C-O, C-S and C-N bonds by means other than hydrolysis or oxidation. Common names include decarboxylase and aldolase. 5. ISOMERASES Catalyze atomic rearrangements within a molecule. Examples include rotamase, protein disulfide isomerase (PDI), epimerase and racemase
, epimerase and racemase.")
35
The role of rotamase and protein disulfide isomerase (PDI)
The role of rotamase and protein disulfide isomerase (PDI). The reactions catalyzed by the two enzymes can assist a peptide chain to fold into a correct three-dimensional structure
. The reactions catalyzed by the two enzymes can assist a peptide chain to fold into a correct three-dimensional structure.")
36
The IUBMB committee also defines subclasses and sub-subclasses
6. LIGASES Catalyze the reaction which joins two molecules Examples include peptide synthase, aminoacyl-tRNA synthetase, DNA ligase and RNA ligase The IUBMB committee also defines subclasses and sub-subclasses Each enzyme is assigned an EC (Enzyme Commission) number For example, the EC number of catalase is EC The first digit indicates that the enzyme belongs to oxidoreductase (class 1) Subsequent digits represent subclasses (1.11. acting on a peroxide as acceptor) and sub-subclasses (1.11.1peroxidases)
 number For example, the EC number of catalase is EC The first digit indicates that the enzyme belongs to oxidoreductase (class 1) Subsequent digits represent subclasses (1.11. acting on a peroxide as acceptor) and sub-subclasses (1.11.1peroxidases)")
37
Mechanism of Enzyme Action
The molecule acted upon a unique geometric shape that is complementary to the geometric shape of a substrate molecule

38
Mechanism of Enzyme Action

39
Mechanism of Enzyme Action
Lock and Key Theory first postulated in 1894 by Emil Fischer The lock is the enzyme and the key is the substrate Only the correctly sized key (substrate) fits into the key hole (active site) of the lock (enzyme)
 fits into the key hole (active site) of the lock (enzyme)")
40
The Lock and Key Hypothesis
Enzyme may be used again Enzyme-substrate complex E S P Reaction coordinate

41
The Induced Fit Theory Postulated by Daniel Koshland
It states that, when substrates approach and bind to an enzyme they induce a conformational change This change is analogous to placing a hand (substrate) into a glove (enzyme)
 into a glove (enzyme)")
42
The Induced Fit Theory Some proteins can change their shape (conformation) When a substrate combines with an enzyme, it induces a change in the enzyme’s conformation The active site is then moulded into a precise conformation Making the chemical environment suitable for the reaction The bonds of the substrate are stretched to make the reaction easier (lowers activation energy)
")
43
The Induced Fit Theory This explains the enzymes that can react with a range of substrates of similar types Hexokinase (a) without (b) with glucose substrate
 without (b) with glucose substrate.")
44
The Induced Fit Theory Assumes that the substrate plays a role in determining the final shape of the enzyme and that the enzyme is partially flexible. This explains why certain compounds can bind to the enzyme but do not react because the enzyme has been distorted too much Other molecules may be too small to induce the proper alignment and therefore cannot react Only the proper substrate is capable of inducing the proper alignment of the active site

45
This is a molecular model of the unbound carboxypeptidase A enzyme
This is a representation of carboxypeptidase A with a substrate (turquoise) bound in the active site. The active site is in the induced conformation. This is a molecular model of the unbound carboxypeptidase A enzyme
 bound in the active site. The active site is in the induced conformation. This is a molecular model of the unbound carboxypeptidase A enzyme.")
46
MECHANISMS TO FACILITATE CATALYSIS
A. CATALYSIS BY PROXIMITY For molecules to react, they must come within bond-forming distance of one another The higher the concentration, the more frequently they will encounter one another and the greater will be their rate of interaction aka entropy reduction ACID-BASE CATALYSIS Can be specific or general “Specific” meaning only protons (H3O+ , specific acid) or OH- ions (specific base)
 or OH- ions (specific base)")
47
MECHANISMS TO FACILITATE CATALYSIS
Proximity: Reaction between bound molecules doesn't require an improbable collision of 2 molecules -- they're already in "contact" (increases the local concentration of reactants). Orientation: Reactants are not only near each other on enzyme, they're oriented in optimal position to react, so the improbability of colliding in correct orientation is taken care of.
. Orientation: Reactants are not only near each other on enzyme, they re oriented in optimal position to react, so the improbability of colliding in correct orientation is taken care of.")
48
Rate enhancement by entropy reduction.
a) bimolecular reaction (high activation energy, low rate)
 bimolecular reaction (high activation energy, low rate)")
49
Rate enhancement by entropy reduction.
b) unimolecular reaction, rate enhanced by factor of 105 due to increased probability of collision/reaction of the 2 groups.
 unimolecular reaction, rate enhanced by factor of 105 due to increased probability of collision/reaction of the 2 groups.")
50
Rate enhancement by entropy reduction.
c) constraint of structure to orient groups better (elimination of freedom of rotation around bonds between reactive groups), rate enhanced by another factor of 103, for 108 total rate enhancement over bimolecular reaction.
 constraint of structure to orient groups better (elimination of freedom of rotation around bonds between reactive groups), rate enhanced by another factor of 103, for 108 total rate enhancement over bimolecular reaction.")
51
MECHANISMS TO FACILITATE CATALYSIS
When substrate binds to enzyme, water is usually excluded from active site (desolvation). causes local dielectric constant to be lower, which enhances electrostatic interactions in the active site, and also results in protection of reactive groups from water, so water doesn't react to form unwanted biproducts. Of course, if water is a substrate, it has to be "allowed in", but maybe only in a certain sub-part of active site. Involvement of charged enzyme functional groups in stabilizing otherwise unstable intermediates in the chemical mechanism can also correctly be called "electrostatic catalysis".
. causes local dielectric constant to be lower, which enhances electrostatic interactions in the active site, and also. results in protection of reactive groups from water, so water doesn t react to form unwanted biproducts. Of course, if water is a substrate, it has to be allowed in , but maybe only in a certain sub-part of active site. Involvement of charged enzyme functional groups in stabilizing otherwise unstable intermediates in the chemical mechanism can also correctly be called electrostatic catalysis .")
52
MECHANISMS TO FACILITATE CATALYSIS
CATALYSIS BY STRAIN Strain is created by binding to substrates in a conformation slightly unfavorable for the bond to undergo cleavage The strain stretches or distorts the targeted bond, weakening it and making it more vulnerable to cleavage probably the most important rate enhancing mechanism available to enzymes Enzyme binds transition state of the reaction more tightly than either the substrate or product -- therefore DG‡ is reduced, and rate is enhanced.

53
Strain "Strain" is a classic concept in which it was supposed that binding of the substrate to the enzyme somehow caused the substrate to become distorted toward the transition state. It's unlikely that there is enough energy available in substrate binding to actually distort the substrate toward the transition state. It's possible that the substrate and enzyme interact unfavorably and this unfavorable interaction is relieved in the transition state. It's more likely that the enzyme is strained, as for example in induced fit.

54
MECHANISMS TO FACILITATE CATALYSIS
Transition state stabilization is a more modern concept: it is not the substrate that is distorted but rather that the transition state makes better contacts with the enzyme than the substrate does, so the full binding energy is not achieved until the transition state is reached. Induced fit assumes that the active site of an enzyme is not complementary to that of the transition state in the absence of the substrate. Such enzymes will have a lower value of kcat/Km, because some of the binding energy must be used to support the conformational change in the enzyme. Induced fit increases Km without increasing kcat.

55
MECHANISMS TO FACILITATE CATALYSIS
COVALENT CATALYSIS Involves the formation of a covalent bond between the enzyme and one or more substrates Introduces a new reaction pathway with lower activation energy thus faster than the reaction pathway in homogenous solution Common among enzymes that catalyze group transfer reactions

56
ENZYME KINETICS The field of biochemistry concerned with the quantitative measurement of the rates of enzyme-catalyzed reactions and the systematic study of factors that affect these rates

57
ENZYME KINETICS REACTION MODEL where S is the substrate
E is the enzyme ES is the enzyme-substrate complex k1, k-1, and k2 are rate constants

58
MICHAELIS MENTEN EQUATION
Describes how reaction velocity varies with substrate concentration Vmax S vo = Km + S where Vo = initial reaction velocity Vmax = maximal velocity Km = Michaelis constant (k-1 + k2)/k1 S= substrate concentration
/k1. S= substrate concentration.")
59
ASSUMPTIONS 1. Relative concentrations of E and S
S >E, so [ES] at any time is small 2. Steady-state assumption [ES] does not change in time E + S = ES = E + P, the rate of formation of ES is equal to that of the breakdown of ES 3. Initial velocity Used in the analysis of enzyme reactions Rate of reaction is measured as soon as E and S are mixed P is very small, the rate of back reaction from P to S can be ignored

60
CONCLUSIONS Characteristics of Km a. Small Km
reflects high affinity of the E for S because a low concentration of S is needed to half-saturate the enzyme – that is, reach a velocity that is ½ Vmax b. Large Km Reflects low affinity of E for S because a high concentration of S is needed to half-saturate the enzyme

61
Effect of substrate concentration on reaction velocities
Small Km for enzyme 1 reflects a high affinity of enzyme for the substrate Large Km for enzyme 2 reflects low affinity of enzyme for the substrate

62
CONCLUSIONS 2. Relationship of velocity to enzyme concentration
The rate of reaction is directly proportional to the enzyme concentration at all substrate concentrations 3. Order of reaction First order - S < Km, the velocity of reaction is roughly proportional to the enzyme concentration Zero order - S > Km, the velocity is constant and equal to Vmax; the rate of reaction is then independent of substrate concentration

63
At high concentration of substrate( [S]>>Km), The velocity of the reaction is zero order – that is, constant and independent OF substrate concentration At At low concentration of substrate( [S]<<Km), The velocity of the reaction is first order – that is, proportional to substrate concentration
![At high concentration of substrate( [S]>>Km), The velocity of the reaction is zero order – that is, constant and independent OF substrate concentration](http://slideplayer.com/slide/6963486/24/images/63/At+high+concentration+of+substrate%28+%5BS%5D%3E%3EKm%29%2C+The+velocity+of+the+reaction+is+zero+order+%E2%80%93+that+is%2C+constant+and+independent+OF+substrate+concentration.jpg "At. At low concentration of substrate( [S]<<Km), The velocity of the reaction is first order – that is, proportional to substrate concentration.")
64
Lineweaver-Burk Plot Also called a double-reciprocal plot
If 1/v0 is plotted VS 1/[S], a straight line is obtained The intercept on the x-axis is equal to -1/Km The intercept on the y-axis is equal to 1/Vmax

65
Lineweaver-Burk Plot Can be used to calculate Km and Vmax as well as to determine the mechanism of enzyme inhibitors Equation describing the Lineweaver-Burk Plot is:

66
INHIBITION OF ENZYME ACTIVITY
INHIBITOR – substance that can diminish the velocity of an enzyme catalyzed reaction TYPES OF INHIBITION: COMPETITIVE INHIBITION NONCOMPETITIVE INHIBITION

67
COMPETITIVE INHIBITION
Inhibitor binds reversibly to the same site that the substrate would normally occupy, and therefore competes with the substrate for that site Inhibitors tend to resemble the structures of a substrate, and thus are termed as substrate analogs

68
COMPETITIVE INHIBITION
Malonate (¯OCOCH2COO¯) competes with Succinate (¯OOCCH2CH2COO¯) for the active site of succinate dehydrogenase (SDH) SDH catalyze the removal of one H atom from each of the 2 methylene C’s of succinate Succinate SDH Malonate SDH
 competes with Succinate (¯OOCCH2CH2COO¯) for the active site of succinate dehydrogenase (SDH) SDH catalyze the removal of one H atom from each of the 2 methylene C’s of succinate. Succinate. SDH. Malonate. SDH.")
69
-2H Fumarate (¯OOC-HC=CH-COO¯) Succinate (¯OOC-CH2-CH2-COO¯) Malonate – Enzyme Complex NO REACTION
 Succinate (¯OOC-CH2-CH2-COO¯) Malonate – Enzyme Complex NO REACTION")
70
Consequences of competitive inhibition
Vmax is unchanged: At high levels of substrate all of the inhibitor is displaced by substrate. Km is increased: Higher substrate concentrations are required to reach the maximal velocity.

71
NONCOMPETITIVE INHIBITION
Inhibitor and substrate bind at different sites on the enzyme The inhibitor binds to both E and ES The noncompetitive inhibitor binds to an allosteric site (different location than the active site) of an enzyme The binding of an inhibitor to the allosteric site alters the shape of the enzyme, resulting in a distorted active site that does not function properly.
 of an enzyme. The binding of an inhibitor to the allosteric site alters the shape of the enzyme, resulting in a distorted active site that does not function properly.")
72
Effect of Enzyme inhibition on Lineweaver-Burk plot

73
NONCOMPETITIVE INHIBITION
Vmax is decreased: At high levels of substrate the inhibitor is still bound. Km is not changed: Noncompetitive inhibitors do not interfere the binding of substrate to enzyme

75
FACTORS AFFECTING ENZYME REACTIONS
I. SUBSTRATE CONCENTRATION The rate of enzyme catalyzed reaction increases with substrate concentration until a maximal velocity (Vmax) is reached
 is reached.")
76
Enzymes are usually damaged
Effect of Temperature The rate of enzyme-catalysed reactions increases as the temperature rises to the optimum temperature Above a certain temperature, activity begins to decline because the enzyme begins to denature Enzymes are usually damaged above about 45°C

77
Effect of pH Each enzyme has an optimal pH
In order to interact, the E and S have specific chemical groups in ionized or unionized state Amino group in protonated form (-NH3+) increase catalytic activity At alkaline pH, amino group is deprotonated decrease in rate of reaction Extremes of pH can lead to denaturation
 increase catalytic activity. At alkaline pH, amino group is deprotonated decrease in rate of reaction. Extremes of pH can lead to denaturation.")
78
REGULATION OF ENZYME ACTIVITY
A. ALLOSTERIC REGULATION B. REGULATION OF ENZYMES BY COVALENT MODIFICATION

79
A. ALLOSTERIC REGULATION
EFFECTORS – molecules that regulate allosteric enzymes that bind noncovalently at a site other than the active site Negative effectors – inhibit enzyme activity Positive effectors – increases enzyme activity

80
HOMOTROPIC EFFECTORS Substrate itself serves as an effector
Most often a positive effector The presence of a substrate molecules at one site on the enzyme enhances the catalytic properties of the other substrate-binding sites(their sites exhibit cooperativity)
")
81
HETEROTROPIC EFFECTORS
The effector may be different from the substrate

82
Feedback Inhibition

83
B. REGULATION OF ENZYMES BY COVALENT MODIFICATION
Most frequently by the addition or removal of phosphate group from specific Ser, Thr, and Tyr residues of the enzyme ATP ADP Protein kinase Enzyme-OPO3= Enzyme-OH Protein phosphatase HPO4= H2O

84
Response of Enzyme to phosphorylation
Phosphorylated form may be more or less active than the unphosphorylated enzyme Glycogen phosphorylase (degrades glycogen) activity is increased low activity (E), high activity (EP) Glycogen synthase (synthesize glycogen) activity is decreased low activity (EP), high activity (E)
 activity is increased. low activity (E), high activity (EP) Glycogen synthase (synthesize glycogen) activity is decreased. low activity (EP), high activity (E)")
85
INDUCTION and REPRESSION of enzyme synthesis
Alter the total population of active sites rather than influencing the efficiency of existing enzyme molecules Enzymes that are needed at only one stage of development or under selected physiologic conditions are subject to regulation of synthesis Enzymes that are in constant use are NOT regulated by altering the rate of enzyme synthesis

86
Mechanisms for Regulating Enzyme Activity
Regulator event Typical effector Results Time required for change Substrate Availability Substrate Change in velocity Immediately Product inhibition Product Change in Vmax and/or Km Allosteric control End product Covalent modification Another enzyme Immediately - minutes Synthesis or degradation of enzyme Hormone or metabolite Change in the amount of enzyme Hours to days

87
Enzyme Activity is Often Regulated
Energy and Enzymes 4/22/2017 Enzyme Activity is Often Regulated Feedback inhibition - a common form of enzyme regulation in which the product inhibits the enzyme . G. Podgorski, Biol. 1010

89
Enzymes - Activity Temperature and pH effect enzyme action

90
Enzymes - Activity Temperature and pH effect enzyme action

91
Enzymes - Activity Enzyme and substrate concentrations

93
ENZYMES IN CLINICAL USE
Enzyme inhibitors as DRUGS Enzymes in CLINICAL DIAGNOSIS

94
Enzyme inhibitors as DRUGS
STATINS – HMG Coenzyme A reductase inhibitors; lower serum lipid concentration EMTRICTABINE and TENOFOVIR DISOPROXIL FUMARATE – inhibitors of viral reverse transcriptase; block replication of HIV ACE Inhibitors (Captopril, Lisinopril, Enalapril) – antihypertensive agents Lactam Antibiotics (Penicillin and Amoxicillin) – inhibitors of alanyl alanine carboxypeptidase-transpeptidase, thus blocking cell wall synthesis
 – antihypertensive agents. Lactam Antibiotics (Penicillin and Amoxicillin) – inhibitors of alanyl alanine carboxypeptidase-transpeptidase, thus blocking cell wall synthesis.")
95
Enzymes in CLINICAL DIAGNOSIS
2 GROUPS OF PLASMA ENZYMES Actively secreted into the plasma by certain organs Released from the cells during normal cell turnover Intracellular, have no physiologic function in the plasma Constant level in healthy individuals and represent a steady state

96
Elevated enzyme activity in the plasma may indicate tissue damage accompanied by increased release of intracellular enzymes, thus useful as a diagnostic tool Elevated levels of ALT (alanine aminotransferase; also called glutamate: pyruvate transaminase; GPT) signals damage
 signals damage.")
97
ISOENZYMES Also called isozymes
Enzymes that catalyze the same reaction but differ in their physical properties because of genetically determined differences in amino acid sequence Different organs frequently contain characteristic proportions of different isoenzymes Isoenzymes found in the plasma serve as a means of identifying the site of tissue damage

98
CK, Creatinine kinase also called Creatinine phosphokinase (CPK)
3 isoenzymes; CK1, CK2, and CK3 Each isoenzyme is a dimer composed of 2 polypeptides (B and M subunits: CK1=BB, CK2=MB, CK3=MM) CK2(MB) isoenzyme is present in more than 5% in myocardial muscles Appears approximately 4 to 8 hours following onset of chest pain, and reaches a peak in activity at approximately 24 hours
 CK2(MB) isoenzyme is present in more than 5% in myocardial muscles. Appears approximately 4 to 8 hours following onset of chest pain, and reaches a peak in activity at approximately 24 hours.")
99
LACTATE DEHYDROGENASE (LDH)
Elevated following an infarction peaking 3 to 6 days after the onset of symptoms Of diagnostic value in patients admitted more than 48 hours after the infarction

100
Principal Serum Enzymes Used in Clinical Diagnosis
Major Diagnostic Use Aminotransferases Aspartate aminotransferase (AST, or SGOT) Alanine aminotransferase (ALT, or SGPT) Myocardial infarction Viral hepatitis Amylase Acute pancreatitis Ceruplasmin Hepatolenticular degeneration (Wilson’s disease) Creatinine kinase Muscle disorders and myocardial infarction
 Alanine aminotransferase (ALT, or SGPT) Myocardial infarction. Viral hepatitis. Amylase. Acute pancreatitis. Ceruplasmin. Hepatolenticular degeneration (Wilson’s disease) Creatinine kinase. Muscle disorders and myocardial infarction.")
101
Principal Serum Enzymes Used in Clinical Diagnosis
Major Diagnostic Use -Glutamyl transpeptidase Various liver diseases Lactate dehydrogenase (isoenzymes) Myocardial infarction Lipase Acute pancreatitis Phosphatase, acid Phosphatase, alkaline Metastatic carcinoma of the prostate Various bone disorders, obstructive liver diseases
 Myocardial infarction. Lipase. Acute pancreatitis. Phosphatase, acid. Phosphatase, alkaline. Metastatic carcinoma of the prostate. Various bone disorders, obstructive liver diseases.")
Similar presentations



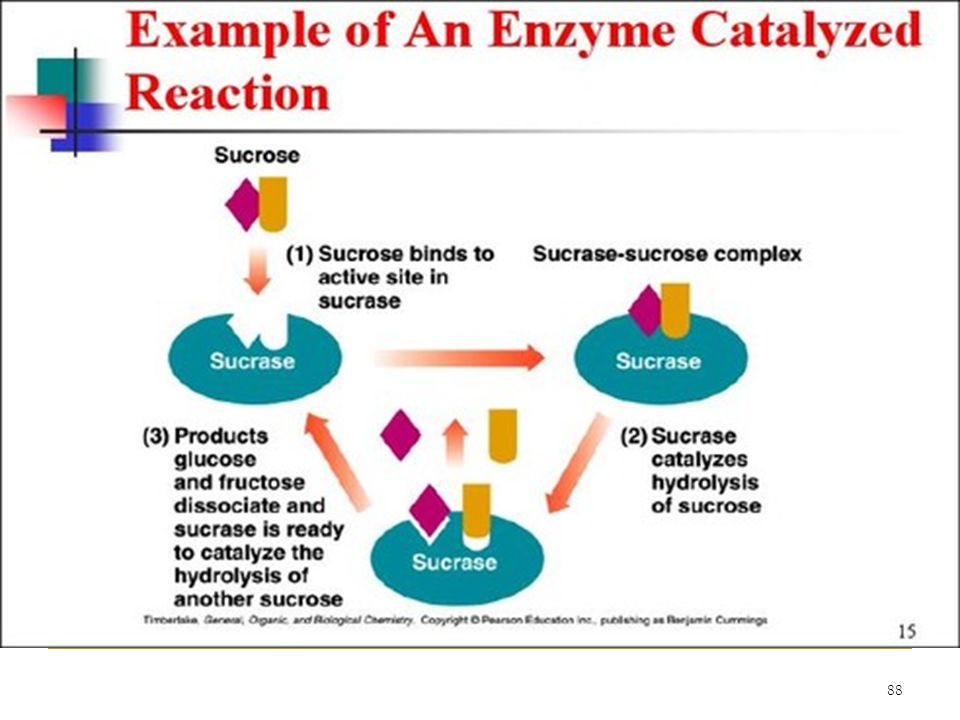









>")






 Dr. Ahmed Sherif Attia https://sites.google.com/site/ahmedsattia/>")



