Download presentation
Presentation is loading. Please wait.

1
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Section C: The Control of Gene Expression 1.Each cell of a multicellular eukaryote expresses only a small fraction of its genes 2. The control of gene expression can occur at any step in the pathway from gene to functional protein: an overview 3. Chromatin modifications affect the availability of genes for transcription 4. Transcription initiation is controlled by proteins that interact with DNA and each other 5. Post-transcriptional mechanisms play supporting roles in the control of gene expression CHAPTER 19 THE ORGANIZATION AND CONTROL OF EUKARYOTIC GENOMES

2
–Like unicellular organisms, the tens of thousands of genes in the cells of multicellular eukaryotes are continually turned on and off in response to signals from their internal and external environments. Gene expression must be controlled on a long-term basis during cellular differentiation, the divergence in form and function as cells specialize. –Highly specialized cells, like nerves or muscles, express only a tiny fraction of their genes. 1. Each cell of a multicellular eukarote expresses only a small fraction of its genes Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

3
Prokaryotic Gene Expression End of Ch 18

4
Regulate-turn on or turn off Genes are either repressible or inducible What would be the initial state of a repressible gene? What would be the initial state of a inducible gene? What would be the road block either to be put in place or to be removed? What would be the signal to turn on or turn off?

5
Students build Prokaryotic Operons See given materials and instructions

6
Repressible Trp operon-when on-it builds Tryptophan

7
Inducible lac operon-when on-it catabolizes allolactose

8
Take 8 Stop Animation Pictures 4 for each gene 1. Take a picture of Gene in its normal state 2. Take a picture of signal that comes in 3.Take a picture of regulatory protein changing shape due to signal adherence 4. Take a picture of gene (operon) in its new state
 in its new state.")
9
An individual bacterium, locked into the genome that it has inherited, can cope with environmental fluctuations by exerting metabolic control. –First, cells vary the number of specific enzyme molecules by regulating gene expression. –Second, cells adjust the activity of enzymes already present (for example, by feedback inhibition). 3. The control of gene expression enables individual bacteria to adjust their metabolism to environmental change Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
. 3. The control of gene expression enables individual bacteria to adjust their metabolism to environmental change Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings.")
10
For example, the tryptophan biosynthesis pathway demonstrates both levels of control. –If tryptophan levels are high, some of the tryptophan molecules can inhibit the first enzyme in the pathway. –If the abundance of tryptophan continues, the cell can stop synthesizing additional enzymes in this pathway by blocking transcription of the genes for these enzymes. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 18.19

11
In 1961, Francois Jacob and Jacques Monod proposed the operon model for the control of gene expression in bacteria. An operon consists of three elements: –the genes that it controls, In bacteria, the genes coding for the enzymes of a particular pathway are clustered together and transcribed (or not) as one long mRNA molecule. –a promotor region where RNA polymerase first binds, –an operator region between the promotor and the first gene which acts as an “on-off switch”. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
 as one long mRNA molecule. –a promotor region where RNA polymerase first binds, –an operator region between the promotor and the first gene which acts as an on-off switch . Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings.")
12
Railroad track analogy

13
By itself, an operon is on and RNA polymerase can bind to the promotor and transcribe the genes. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 18.20a

14
However, if a repressor protein, a product of a regulatory gene, binds to the operator, it can prevent transcription of the operon’s genes. –Each repressor protein recognizes and binds only to the operator of a certain operon. –Regulatory genes are transcribed at low rates continuously. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 18.20b

16
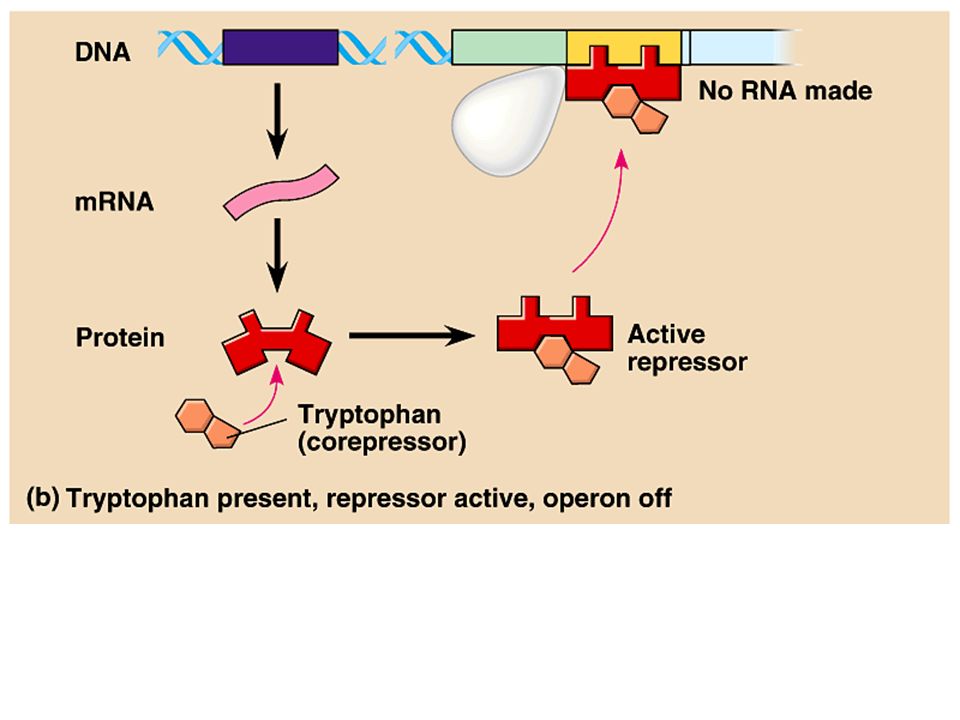
Binding by the repressor to the operator is reversible. –The number of active repressor molecules available determines the on and off mode of the operator. Many repressors contain allosteric sites that change shape depending on the binding of other molecules. –In the case of the trp operon, when concentrations of tryptophan in the cell are high, some tryptophan molecules bind as a corepressor to the repressor protein. –This activates the repressor and turns the operon off. –At low levels of tryptophan, most of the repressors are inactive and the operon is transcribed. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

17
The trp operon is an example of a repressible operon, one that is inhibited when a specific small molecule binds allosterically to a regulatory protein. In contrast, an inducible operon is stimulated when a specific small molecule interacts with a regulatory protein. –In inducible operons, the regulatory protein is active (inhibitory) as synthesized, and the operon is off. –Allosteric binding by an inducer molecule makes the regulatory protein inactive, and the operon is on. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
 as synthesized, and the operon is off. –Allosteric binding by an inducer molecule makes the regulatory protein inactive, and the operon is on. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings.")
18
The lac operon, containing a series of genes that code for enzymes, which play a major role is the hydrolysis and metabolism for lactose. –In the absence of lactose, this operon is off as an active repressor binds to the operator and prevents transcription. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 18.21a

20
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 18.21b When lactose is present in the cell, allolactase, an isomer of lactose, binds to the repressor. This inactivates the repressor, and the lac operon can be transcribed.

21
Repressible enzymes generally function in anabolic pathways, synthesizing end products. –When the end product is present in sufficient quantities, the cell can allocate its resources to other uses. Inducible enzymes usually function in catabolic pathways, digesting nutrients to simpler molecules. –By producing the appropriate enzymes only when the nutrient is available, the cell avoids making proteins that have nothing to do. Both repressible and inducible operons demonstrate negative control because active repressors can only have negative effects on transcription. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

22
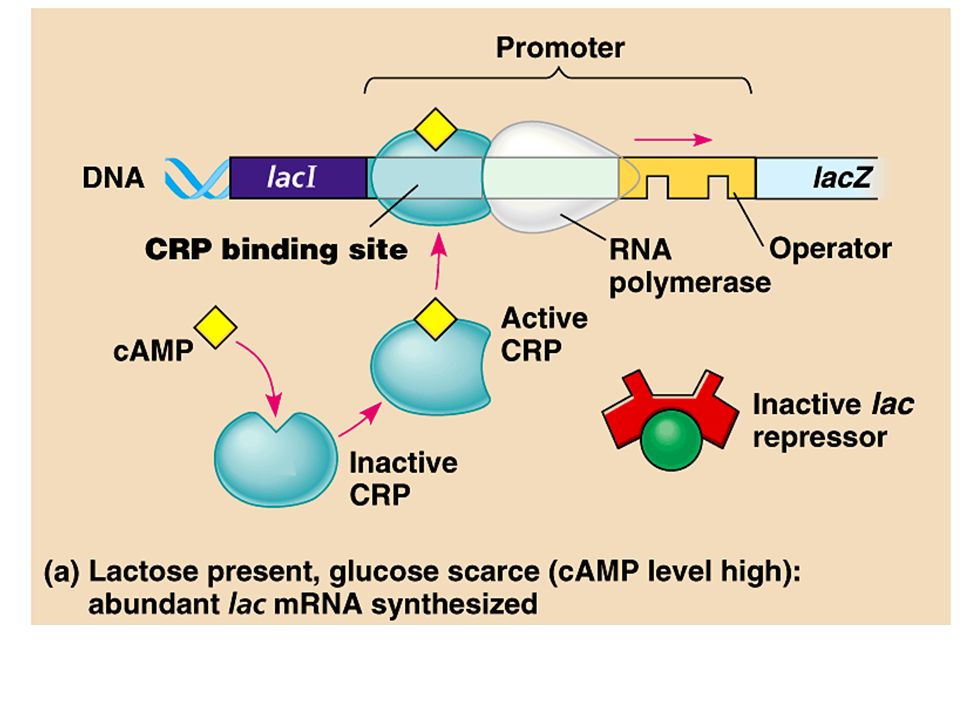
Positive gene control occurs when an activator molecule interacts directly with the genome to switch transcription on. Even if the lac operon is turned on by the presence of allolactose, the degree of transcription depends on the concentrations of other substrates. –If glucose levels are low (along with overall energy levels), then cyclic AMP (cAMP) binds to cAMP receptor protein (CRP) which activates transcription. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 18.22a
, then cyclic AMP (cAMP) binds to cAMP receptor protein (CRP) which activates transcription. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig a.")
24
The cellular metabolism is biased toward the utilization of glucose. If glucose levels are sufficient and cAMP levels are low (lots of ATP), then the CRP protein has an inactive shape and cannot bind upstream of the lac promotor. –The lac operon will be transcribed but at a low level. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 18.22b
, then the CRP protein has an inactive shape and cannot bind upstream of the lac promotor. –The lac operon will be transcribed but at a low level. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig b.")
25
For the lac operon, the presence / absence of lactose (allolactose) determines if the operon is on or off. Overall energy levels in the cell determine the level of transcription, a “volume” control, through CRP. CRP works on several operons that encode enzymes used in catabolic pathways. –If glucose is present and CRP is inactive, then the synthesis of enzymes that catabolize other compounds is slowed. –If glucose levels are low and CRP is active, then the genes which produce enzymes that catabolize whichever other fuel is present will be transcribed at high levels. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

26
Free Response over Trxn Regulation

27
Peer Grading

28
Part II

29
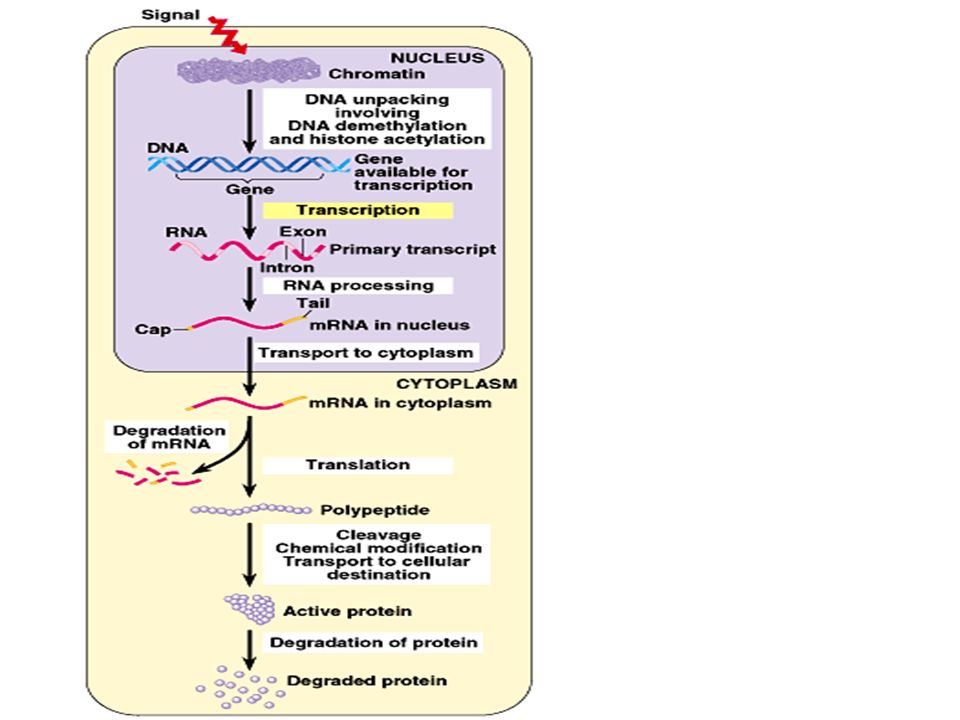
Each stage in the entire process of gene expression provides a potential control point where gene expression can be turned on or off, speeded up or slowed down. A web of control connects different genes and their products. 2. The control of gene expression can occur at any step in the pathway from gene to functional proteins: an overview Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

30
Eukaryotic gene expression Big Toe question How do you think eukaryotic gene regulation is different/similar to the prokaryotic operon system?

31
Controls of gene activity in eukaryotes involves some of the principles described for prokaryotes. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

32
–The expression of specific genes is commonly regulated at the transcription level by DNA- binding proteins that interact with other proteins and with external signals. –WHAT WERE THE NAMES OF THESE BINDING PROTEINS? –With their greater complexity, eukaryotes have opportunities for controlling gene expression at additional stages.

33
These levels of control include : chromatin packing, transcription, RNA processing, translation, and various alterations to the protein product. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 19.7

35
PACKAGING Affects availability of a gene to even receive a signal video

36
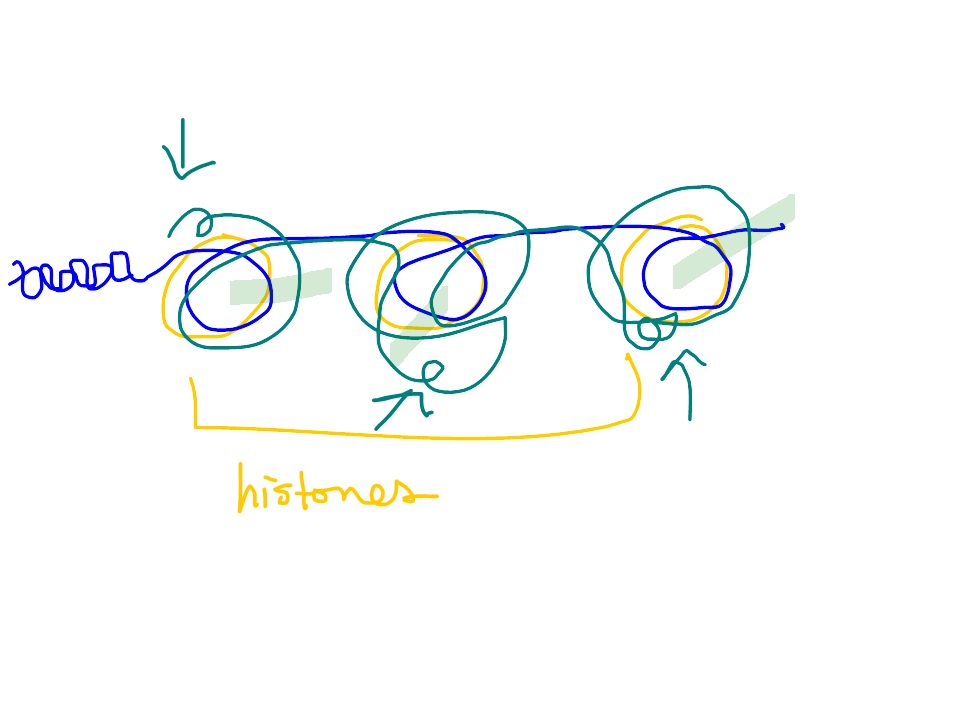
In addition to its role in packing DNA inside the nucleus, chromatin organization impacts regulation. –Genes of densely condensed heterochromatin are usually not expressed, presumably because transcription proteins cannot reach the DNA. –A gene’s location relative to nucleosomes and to attachments sites to the chromosome scaffold or nuclear lamina can affect transcription. Chromatin modifications affect the availability of genes for transcription Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

37
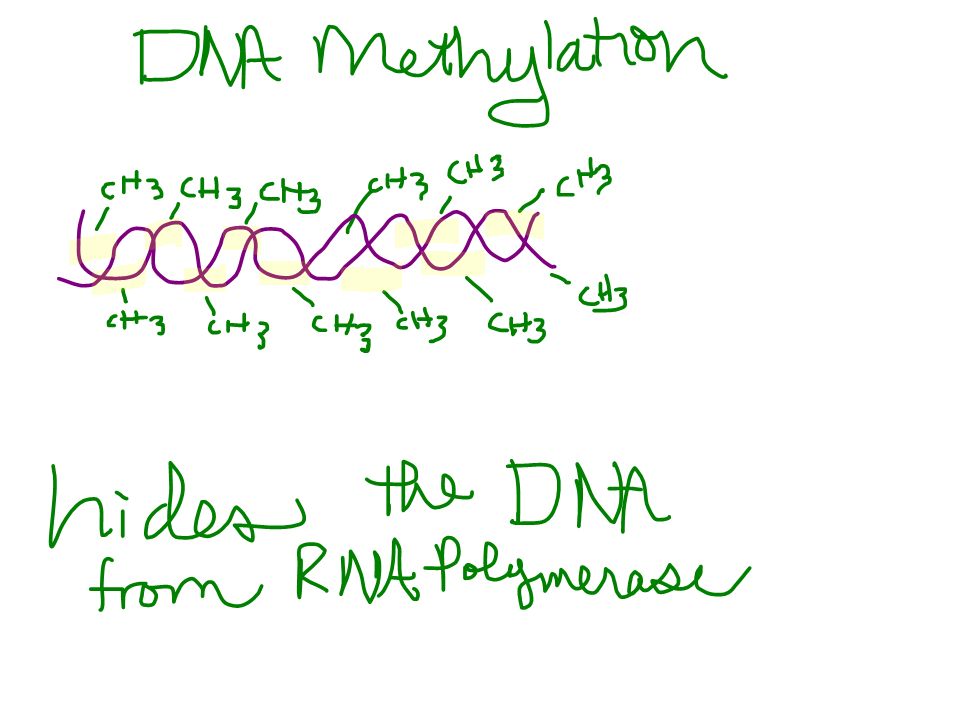
DNA methylation is the attachment by specific enzymes of methyl groups (-CH 3 ) to DNA bases after DNA synthesis. Inactive DNA is generally highly methylated compared to DNA that is actively transcribed. –For example, the inactivated mammalian X chromosome in females is heavily methylated. –Genes are usually more heavily methylated in cells where they are not expressed. –Demethylating certain inactive genes turns them on. –This process was used to turn skin cells back into noncontroversial stem cells Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

39
In some species DNA methylation is responsible for long-term inactivation of genes during cellular differentiation. –Once methylated, genes usually stay that way through successive cell divisions. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

40
–Methylation enzymes recognize sites on one strand that are already methylated and correctly methylate the daughter strand after each round of DNA replication. –This methylation patterns accounts for genomic imprinting in which methylation turns off either the maternal or paternal alleles of certain genes at the start of development.

41
Prader Willi versus Angelman Syndrome

42
For most genes it is a reasonable assumption that a specific allele will have the same effect regardless of whether it was inherited from the mother or father. However, for some traits in mammals, it does depend on which parent passed along the alleles for those traits. –The genes involved are not sex linked and may or may not lie on the X chromosome. 2. The phenotypic effects of some mammalian genes depend on whether they were inherited from the mother or the father (imprinting) Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
 Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings.")
43
Two disorders, Prader-Willi syndrome and Angelman syndrome, with different phenotypic effects are due to the same cause, a deletion of a specific segment of chromosome 15. –Individuals with Prader-Willi syndrome are characterized by mental retardation, obesity, short stature, and unusually small hands and feet. –These individuals inherit the abnormal chromosome from their father. –Individuals with Angelman syndrome exhibit spontaneous laughter, jerky movements, and other motor and mental symptoms. –This is inherited from the mother. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

44
The difference between the disorders is due to genomic imprinting. In this process, a gene on one homologous chromosome is silenced, while its allele on the homologous chromosome is expressed. The imprinting status of a given gene depends on whether the gene resides in a female or a male. –The same alleles may have different effects on offspring, depending on whether they arrive in the zygote via the ovum or via the sperm. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

45
In the new generation, both maternal and paternal imprints are apparently “erased” in gamete-producing cells. Then, all chromosomes are reimprinted according to the sex of the individual in which they reside. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 15.15

46
In many cases, genomic imprinting occurs when methyl groups are added to cytosine nucleotides on one of the alleles. –Heavily methylated genes are usually inactive. –The animal uses the allele that is not imprinted. In other cases, the absence of methylation in the vicinity of a gene plays a role in silencing it. –The active allele has some methylation. Several hundred mammalian genes, many critical for development, may be subject to imprinting. –Imprinting is critical for normal development. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

47
Fragile X syndrome, which leads to various degrees of mental retardation, also appears to be subject to genomic imprinting. –This disorder is named for an abnormal X chromosome in which the tip hangs on by a thin thread of DNA. –This disorder affects one in every 1,500 males and one in every 2,500 females. Inheritance of fragile X is complex, but the syndrome is more common when the abnormal chromosome is inherited from the mother. –This is consistent with the higher frequency in males. –Imprinting by the mother somehow causes it. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

48
Histone acetylation (addition of an acetyl group -COCH 3 ) and deacetylation appear to play a direct role in the regulation of gene transcription. –Acetylated histones grip DNA less tightly, providing easier access for transcription proteins in this region. –Some of the enzymes responsible for acetylation or deacetylation are associated with or are components of transcription factors that bind to promoters. –In addition, some DNA methylation proteins recruit histone deacetylation enzymes, providing a mechanism by which DNA methylation and histone deacetylation cooperate to repress transcription. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

50
–Chromatin-modifying enzymes provide a coarse adjustment to gene expression by making a region of DNA either more available or less available for transcription. Fine-tuning begins with the interaction of transcription factors with DNA sequences that control specific genes. Initiation of transcription is the most important and universally used control point in gene expression. 4. Transcription initiation is controlled by proteins that interact with DNA and with each other Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

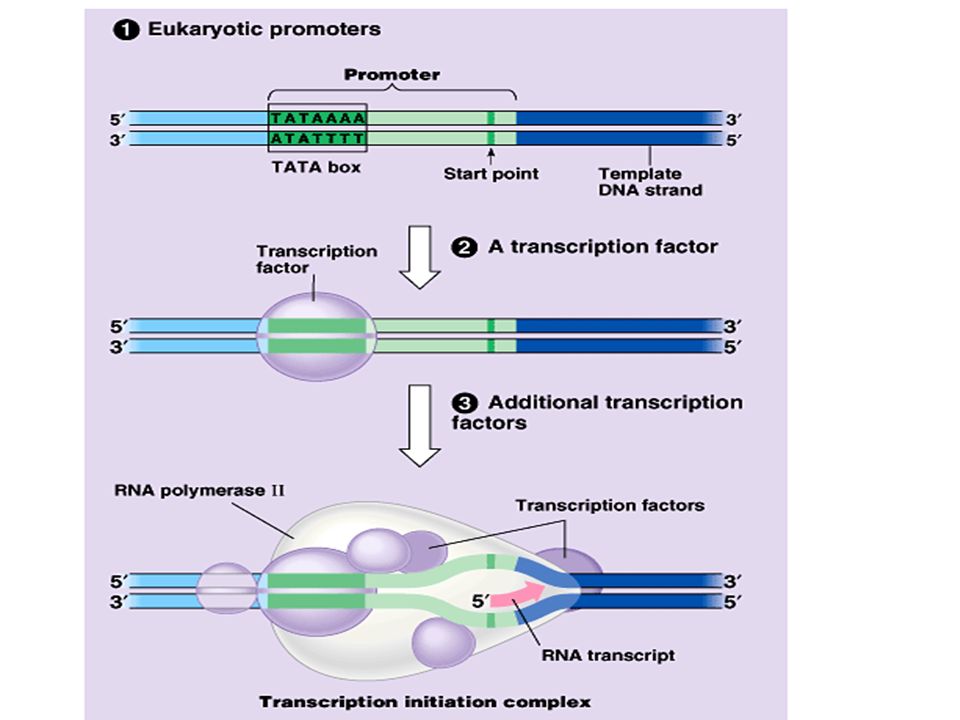
51
A eukaryotic gene and the DNA segments that control transcription include introns and exons, a promoter sequence upstream of the gene, and a large number of other control elements. –Control elements are noncoding DNA segments that regulate transcription by binding transcription factors. –The transcription initiation complex assembles on the promotor sequence and one component, RNA polymerase, then transcribes the gene. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

52
Fig. 19.8

53
Eukaryotic RNA polymerase is dependent on transcription factors before transcription begins. –One transcription factor recognizes the TATA box. –Others in the initiation complex are involved in protein-protein interactions. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

55
High transcription levels require additional transcription factors binding to other control elements. Distant control elements, enhancers, may be thousands of nucleotides away from the promoter or even downstream of the gene or within an intron.

56
Bending of DNA enables transcription factors, activators, bound to enhancers to contact the protein initiation complex at the promoter. –This helps position the initiation complex on the promoter. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 19.9

57
Eukaryotic genes also have repressor proteins that bind to DNA control elements called silencers. At the transcription level, activators are probably more important than repressors, because the main regulatory mode of eukaryotic cells seems to be activation of otherwise silent genes. Repression may operate mostly at the level of chromatin modification. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

58
The hundreds of eukaryotic transcription factors follow only a few basic structural principles. –Each protein generally has a DNA-binding domain that binds to DNA and a protein- binding domain that recognizes other transcription factors. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 19.10

59
A surprisingly small number of completely different nucleotide sequences are found in DNA control elements. Members of a dozen or so sequences about four to ten base pairs long appear again and again in the control elements for different genes. For many genes, the particular combination of control elements associated with the gene may be more important than the presence of a control element unique to the gene. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

60
In contrast, only rarely are eukaryotic genes organized this way. –Genes coding for the enzymes of a metabolic pathway may be scattered over different chromosomes. –Even if genes are on the same chromosome, each gene has its own promoter and is individually transcribed. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

61
In prokaryotes, coordinately controlled genes are often clustered into an operon with a single promoter and other control elements upstream. –The genes of the operon are transcribed into a single mRNA and translated together

62
Coordinate gene expression in eukaryotes probably depends on the association of a specific control element or collection of control elements with every gene of a dispersed group. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

63
A common group of transcription factors bind to them, promoting simultaneous gene transcription. –For example, steroid hormones enter a cell and bind to a specific receptor protein in the cytoplasm or nucleus. –After allosteric activation of these proteins, they functions as transcription activators. –Other signal molecules can control gene expression indirectly by triggering signal-transduction pathways that lead to transcription activators.

64
Gene expression may be blocked or stimulated by any post-transcriptional step. By using regulatory mechanisms that operate after transcription, a cell can rapidly fine-tune gene expression in response to environmental changes without altering its transcriptional patterns. 5. Post-transcriptional mechanisms pay supporting roles in the control of gene expression Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

65
RNA processing in the nucleus and the export of mRNA to the cytoplasm provide opportunities for gene regulation that are not available in bacteria. In alternative RNA splicing, different mRNA molecules are produced from the same primary transcript, depending on which RNA segments are treated as exons and which as introns. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 19.11

66
The life span of a mRNA molecule is an important factor determining the pattern of protein synthesis. Prokaryotic mRNA molecules may be degraded after only a few minutes. Eukaryotic mRNAs endure typically for hours or even days or weeks. –For example, in red blood cells the mRNAs for the hemoglobin polypeptides are unusually stable and are translated repeatedly in these cells. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

67
A common pathway of mRNA breakdown begins with enzymatic shortening of the poly(A) tail. –This triggers the enzymatic removal of the 5’ cap. –This is followed by rapid degradation of the mRNA by nucleases. Nucleotide sequences in the untranslated trailer region at the 3’ end affect mRNA stability. –Transferring such a sequence from a short- lived mRNA to a stable mRNA results in quick mRNA degradation. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

68
Translation of specific mRNAs can be blocked by regulatory proteins that bind to specific sequences or structures within the 5’ leader region of mRNA. –This prevents attachment to ribosomes. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

69
Protein factors required to initiate translation in eukaryotes offer targets for simultaneously controlling translation of all the mRNA in a cell. –This allows the cell to shut down translation if environmental conditions are poor (for example, shortage of a key constituent) or until the appropriate conditions exist (for example, until after fertilization or during daylight in plants).
 or until the appropriate conditions exist (for example, until after fertilization or during daylight in plants)..")
70
Finally, eukaryotic polypeptides must often be processed to yield functional proteins. –This may include cleavage, chemical modifications, and transport to the appropriate destination. Regulation may occur at any of these steps. –For example, cystic fibrosis results from mutations in the genes for a chloride ion channel protein that prevents it from reaching the plasma membrane. –The defective protein is rapidly degraded. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

71
The cell limits the lifetimes of normal proteins by selective degradation. –Many proteins, like the cyclins in the cell cycle, must be short-lived to function appropriately. Proteins intended for degradation are marked by the attachment of ubiquitin proteins. Giant proteosomes recognize the ubiquitin and degrade the tagged protein. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 19.12

73
So-what do you remember? How are genes regulated in prokaryotes?

74
So-what do you remember? How are genes regulated in eukaryotes?

75
Programmed Cellular Death Apoptosis

76
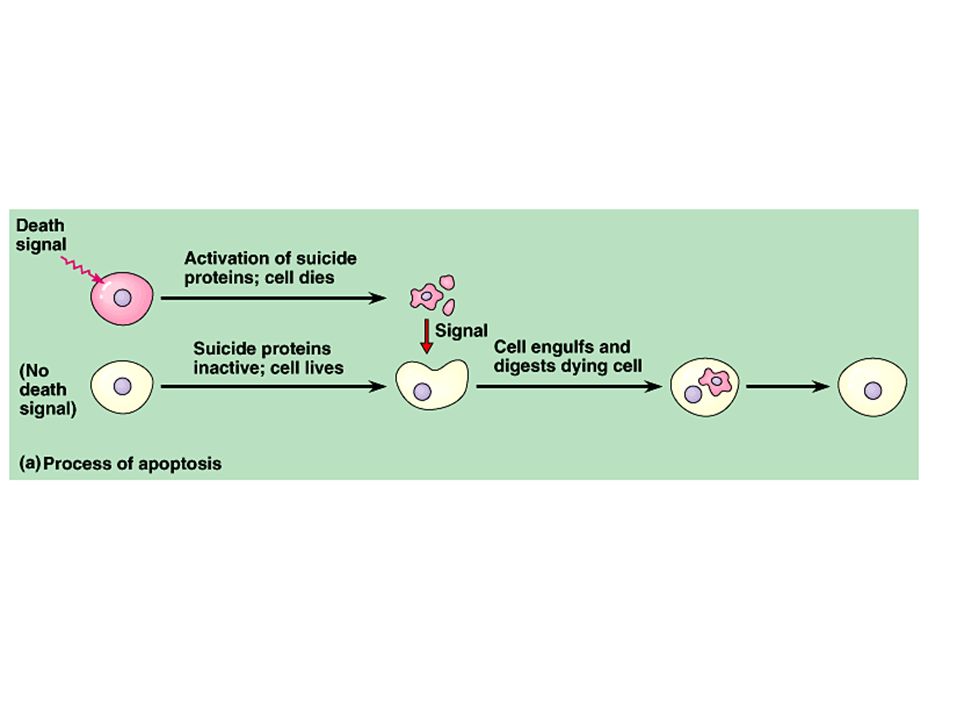
Lineage analysis of C. elegans highlights another outcome of cell signaling, programmed cell death or apoptosis. –The timely suicide of cells occurs exactly 131 times in the course of C. elegans’s normal development. –At precisely the same points in development, signals trigger the activation of a cascade of “suicide” proteins in the cells destined to die. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 21.18a

78
A cell remains alive as long as the Ced-9 protein, produced by the ced-9 gene (ced stands for cell death) is active. –Ced-9, the master regulator of apoptosis, blocks the activation of Ced-4 (produced by ced-4) preventing it from activating Ced-3 (produced by ced-3), a potent protease. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
 preventing it from activating Ced-3 (produced by ced-3), a potent protease. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings.")
79
When the cell receives an external death signal, Ced-9 is inactivated, allowing both Ced-4 and Ced-3 to be active. –In nematodes Ced-3 is the chief caspase, the main proteases of apoptosis.

80
Apoptosis is regulated not at the level of transcription or translation, but through changes in the activity of proteins that are continually present in the cell. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 21.18b

81
Using nematodes as a model

82
Apoptosis pathways in humans and other mammals are more complicated. Research on mammals have revealed a prominent role for mitochondria in apoptosis. –Signals from apoptosis pathways or others somehow cause the outer mitochondrial membrane to leak, releasing proteins that promote apoptosis. –Still controversial is whether mitochondria play a central role in apoptosis or only a subsidiary role. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

83
A cell must make a life-or-death “decision” by somehow integrating both the “death” and “life” (growth factor) signals that it receives.
 signals that it receives.")
84
A built-in cell suicide mechanism is essential to development in all animals. –Similarities between the apoptosis genes in mammals and nematodes indicate that the basic mechanism evolved early in animal evolution. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

85
–The timely activation of apoptosis proteins in some cells functions during normal development and growth in both embryos and adults. It is part of the normal development of the nervous system, normal operation of the immune system, and for normal morphogenesis of human hands and feet.

86
Problems with the cell suicide mechanism may have health consequences, ranging from minor to serious. –Failure of normal cell death during morphogenesis of the hands and feet can result in webbed fingers and toes. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

87
–Researchers are also investigating the possibility that certain degenerative diseases of the nervous system result from inappropriate activation of the apoptosis genes. –Others are investigating the possibility that some cancers result from a failure of cell suicide which normally occurs if the cell has suffered irreparable damage, especially DNA damage.

88
what do you know

89
Cell Death Free Response

90
Rubric Continued

91
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Section D: The Molecular Biology of Cancer 1.Cancer results from genetic changes that affect the cell cycle 2. Oncogene proteins and faulty tumor-suppressor proteins interfere with normal signaling pathways 3. Multiple mutations underlie the development of cancer

92
WHAT ARE ONCOGENES?

93
Cancer is a disease in which cells escape from the control methods that normally regulate cell growth and division. The agent of such changes can be random spontaneous mutations or environmental influences such as chemical carcinogens or physical mutagens. 1. Cancer results from genetic changes that affect the cell cycle Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

94
Cancer-causing genes, oncogenes, were initially discovered in retroviruses, but close counterparts, proto-oncogenes were found in other organisms.

95
The products of proto-oncogenes, proteins that stimulate normal cell growth and division, have essential functions in normal cells. An oncogene arises from a genetic change that leads to an increase in the proto- oncogene’s protein or the activity of each protein molecule. These genetic changes include movements of DNA within the genome, amplification of proto-oncogenes, and point mutations in the gene. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

96
Malignant cells frequently have chromosomes that have been broken and rejoined incorrectly. –This may translocate a fragment to a location near an active promotor or other control element. Amplification increases the number of gene copies. A point mutation may lead to translation of a protein that is more active or longer-lived. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Malignant vs. Benign

97
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 19.13

98
Mutations to genes whose normal products inhibit cell division, tumor-suppressor genes, also contribute to cancer. Any decrease in the normal activity of a tumor- suppressor protein may contribute to cancer. –Some tumor-suppressor proteins normally repair damaged DNA, preventing the accumulation of cancer-causing mutations. –Others control the adhesion of cells to each other or to an extracellular matrix, crucial for normal tissues. –Still others are components of cell-signaling pathways that inhibit the cell cycle. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

99
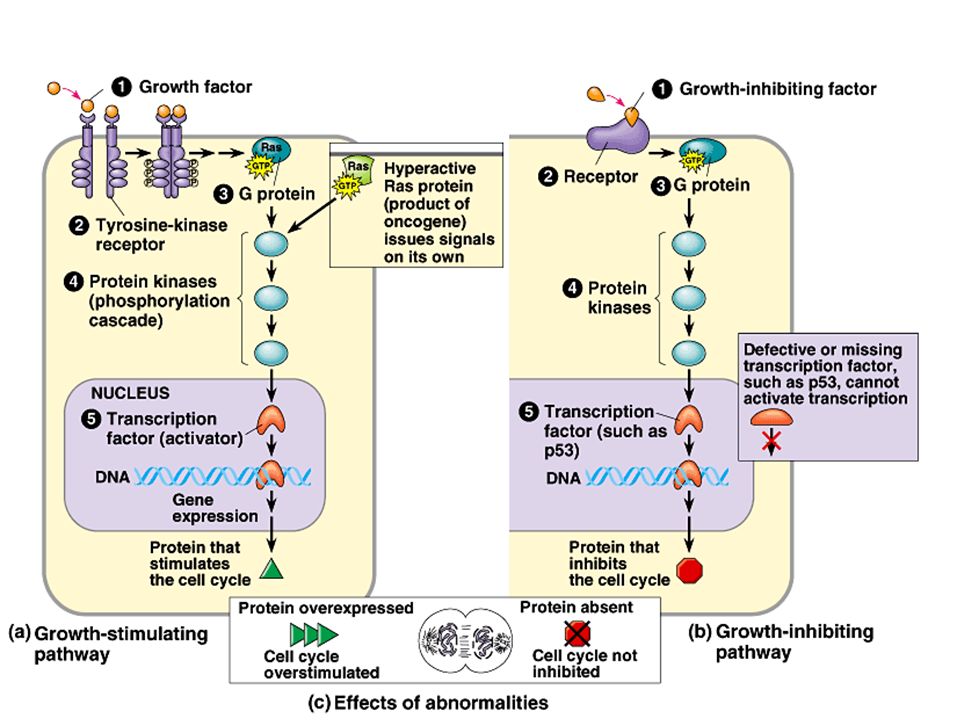
Mutations in the products of two key genes, the ras proto-oncogene, and the p53 tumor suppressor gene occur in 30% and 50% of human cancers respectively. Both the Ras protein and the p53 protein are components of signal-transduction pathways that convey external signals to the DNA in the cell’s nucleus. 2. Oncogene proteins and faulty tumor- suppressor proteins interfere with normal signaling pathways Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

100
Ras, the product of the ras gene, is a G protein that relays a growth signal from a growth factor receptor to a cascade of protein kinases. –At the end of the pathway is the synthesis of a protein that stimulates the cell cycle. –Many ras oncogenes have a point mutation that leads to a hyperactive version of the Ras protein that can issue signals on its own, resulting in excessive cell division. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

101
Do we have an evolutionary adaption to these oncogenes?

102
The tumor-suppressor protein encoded by the normal p53 gene is a transcription factor that promotes synthesis of growth- inhibiting proteins. –A mutation that knocks out the p53 gene can lead to excessive cell growth and cancer.

103
Mutations that result in stimulation of growth- stimulating pathways or deficiencies in growth- inhibiting pathways lead to increased cell division. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 19.14

105
The p53 gene, named for its 53,000-dalton protein product, is often called the “guardian angel of the genome”. Damage to the cell’s DNA acts as a signal that leads to expression of the p53 gene. The p53 protein is a transcription factor for several genes. –It can activate the p21 gene, which halts the cell cycle. –It can turn on genes involved in DNA repair. –When DNA damage is irreparable, the p53 protein can activate “suicide genes” whose protein products cause cell death by apoptosis. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

106
More than one somatic mutation is generally needed to produce the changes characteristic of a full-fledged cancer cell. If cancer results from an accumulation of mutations, and if mutations occur throughout life, then the longer we live, the more likely we are to develop cancer. 3. Multiple mutations underlie the development of cancer Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

107
Colorectal cancer, with 135,000 new cases in the U.S. each year, illustrates a multi- step cancer path. The first sign is often a polyp, a small benign growth in the colon lining with fast dividing cells. Through gradual accumulation of mutations that activate oncogenes and knock out tumor-suppressor genes, the polyp can develop into a malignant tumor. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

108
Fig. 19.15

109
About a half dozen DNA changes must occur for a cell to become fully cancerous. These usually include the appearance of at least one active oncogene and the mutation or loss of several tumor-suppressor genes. –Since mutant tumor-suppressor alleles are usually recessive, mutations must knock out both alleles. –Most oncogenes behave as dominant alleles. In many malignant tumors, the gene for telomerase is activated, removing a natural limit on the number of times the cell can divide. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

110
Viruses, especially retroviruses, play a role is about 15% of human cancer cases worldwide. –These include some types of leukemia, liver cancer, and cancer of the cervix. Viruses promote cancer development by integrating their DNA into that of infected cells. By this process, a retrovirus may donate an oncogene to the cell. Alternatively, insertion of viral DNA may disrupt a tumor-suppressor gene or convert a proto-oncogene to an oncogene. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

111
The fact that multiple genetic changes are required to produce a cancer cell helps explain the predispositions to cancer that run in some families. –An individual inheriting an oncogene or a mutant allele of a tumor-suppressor gene will be one step closer to accumulating the necessary mutations for cancer to develop. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

112
Geneticists are devoting much effort to finding inherited cancer alleles so that predisposition to certain cancers can be detected early in life. –About 15% of colorectal cancers involve inherited mutations, especially to DNA repair genes or to the tumor-suppressor gene APC. Normal functions of the APC gene include regulation of cell migration and adhesion. –Between 5-10% of breast cancer cases, the 2 nd most common U.S. cancer, show an inherited predisposition. Mutations to one of two tumor-suppressor genes, BRCA1 and BRCA2, increases the risk of breast and ovarian cancer. Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

113
Cancer Timeline Create a timeline from beginning of life to tumor creation within a patient. Pick one of the following scenarios: A patient that inherited an oncogene or mutated suppressor gene OR A patient that got a mutation either by carcinogen or mutated DNA caused by a virus

114
Within this timeline… Label the timeline with a sequence of imaginary events that would have led to cancer developing in the patient. Make sure that you include the following details: a. proto-oncogenes that transpose to an actively promoted site

Similar presentations

















. Gene.>")



