Download presentation
Presentation is loading. Please wait.

1
Part Ⅲ Expression of the Gene Chapter 13 RNA Splicing

2
OUTLINE Abstract Contents The Chemistry of RNA Splicing The Spliceosome Machinery Splicing Pathways Alternative Splicing Exon Shuffling RNA Editing mRNA Transport Summary

3
Abstract

4
The constructive ways of the coding sequence: 1. In the vast majority of cases in bacteria and their phage: the coding sequence is contiguous, the codon for one amino acid is immediataly adjacent to the codon for the next amino acid in the polypeptide. 2. In eukaryotic genes: the coding sequence is periodically interrupted by stretches of non-coding sequence.

5
In eukaryotic genes ● Coding sequences: exons ● Intervening sequences: introns —— “mosaics” As a consequence of this alternating pattern of exons and introns, genes bearing non-coding interruptions are often said to be “in pieces” or “split”.

6
Primary transcript

7
Introns 1. Number varies enormously. E.g. One in the case of most intron-containing yeast genes and a few human genes. 50 in the case of the chicken proα2 collagen gene. 363 in the case of the Titin gene of humans. 2. The sizes of the exons and introns vary—— introns are very often much longer than the exons they separate. E.g. ① 150 nucleotides in exons: 800kb in introns. ② The mammalian gene for the enzyme dihydrofolate reductase: 6 exons correspond to 2kb of mRNA: ( ≧ ) 31kb long of the whole genes [i.e. the coding portion of the gene is less than 10% of its total length.]
 31kb long of the whole genes [i.e. the coding portion of the gene is less than 10% of its total length.].")
8
● RNA splicing: the process by which introns are removed from the pre-mRNA. ● Alternative splicing: some pre-mRNAs can be spliced in more than one way, generating alternative mRNAs. 60% of the human genes are spliced in this manner.

9
Topic Ⅰ The Chemistry of RNA Splicing

10
Ⅰ Sequences within the RNA Determine Where Splicing Occurs The borders between introns and exons are marked by specific nucleotide sequences within the pre- mRNAs.

12
● 5’splice site: the exon-intron boundary at the 5’ end of the intron ● 3’ splice site: the exon-intron boundary at the 3’ end of the intron ● Branch point site: an A close to the 3’ end of the intron, which is followed by a polypyrimidine tract (Py tract).
.")
13
Ⅱ The intron is removed in a Form Called a Lariat as the Flanking Exons are joined Two successive transesterification Step 1Step 1: The OH of the conserved A at the branch site attacks the phosphoryl group of the conserved G in the 5’ splice site. As a result, the 5’ exon is released and the 5’-end of the intron forms a three-way junction structure. Step 2Step 2: The OH of the 5’ exon attacks the phosphoryl group at the 3’ splice site. As a consequence, the 5’ and 3’ exons are joined and the intron is liberated in the shape of a lariat.

14
Three-way junction

15
The structure of three-way function

17
Ⅲ Exons from different RNA molecules can be fused by Trans-splicing Trans-splicing: the process in which two exons carried on different RNA molecules can be spliced together.

18
Trans-splicing Not a lariat

19
Topic Ⅱ The Spliceosome Machinery

20
RNA splicing is carried out by a large complex called spliceosome ● The above described splicing of introns from pre-mRNA are mediated by the spliceosome. ● The spliceosome comprises about 150 proteins and 5 snRNAs. ● Many functions of the spliceosome are carried out by its RNA components.

21
● The five RNAs (U1, U2, U4, U5, and U6, 100-300 nt) are called small nuclear RNAs (snRNAs). ● The complexes of snRNA and proteins are called small nuclear ribonuclear proteins (snRNP, pronounces “snurps”). ● The spliceosome is the largest snRNP, and the exact makeup differs at different stages of the splicing reaction.
. ● The spliceosome is the largest snRNP, and the exact makeup differs at different stages of the splicing reaction..")
22
Three roles of snRNPs in splicing 1. Recognizing the 5’ splice site and the branch site. 2. Bringing those sites together. 3. Catalyzing (or helping to catalyze) the RNA cleavage. RNA-RNA, RNA-protein and protein-protein interactions are all important during splicing.
 the RNA cleavage. RNA-RNA, RNA-protein and protein-protein interactions are all important during splicing..")
23
RNA-RNA interactions between different snRNPs, and between snRNPs and pre-mRNA

24
Topic Ⅲ The Spliceosome Machinery

25
Ⅰ Assembly, rearrangement, and catalysis within the spliceosome: the splicing pathway Assembly step 1 1. U1 recognize 5’ splice site. 2. One subunit of U2AF binds to Py tract and the other to the 3’ splice site. The former subunits interacts with BBP and helps it bind to the branch point. 3. Early (E) complex is formed
 complex is formed.")
26
Assembly step 2 1. U2 binds to the branch site, and then A complex is formed. 2. The base-pairing between the U2 and the branch site is such that the branch site A is extruded. This A residue is available to react with the 5’ splice site.

27
E complex A complex

28
Assembly step 3 1. U4, U5 and U6 form the tri-snRNP Particle. 2. With the entry of the tri-snRNP, the A complex is converted into the B complex.

29
A complex B complex

30
C complex in which the catalysis has not occurred yet

31
Catalysis Step 1 ● Formation of the C complex produces the active site, with U2 and U6 RNAs being brought together. ● Formation of the active site juxtaposes the 5’ splice site of the pre-mRNA and the branch site, allowing the branched A residue to attack the 5’ splice site to accomplish the first transesterfication reaction.

32
Catalysis Step 2 U5 snRNP helps to bring the two exons together, and aids the second transesterification reaction, in which the 3’-OH of the 5’ exon attacks the 3’ splice site. Final Step Release of the mRNA product and the snRNPs.

33
C complex

34
E complex A complex B complex C complex splicesome-mediated splicing reactions

35
Ⅱ Self-splicing introns reveal that RNA can catalyze RNA splicing Self-splicing introns: the intron itself folds into a specific conformation within the precursor RNA and catalyzes the chemistry of its own release and the exon ligation.

36
Adams et al., Nature 2004, Crystal structure of a self- splicing group I intron with both exons

37
● Practical definition for self-splicing introns: the introns that can remove themselves from pre-RNAs in the test tube in the absence of any proteins or other RNAs. ● There are two classes of self-splicing introns, group I and group II self-splicing introns.

38
Three class of RNA Splicing ClassAbundanceMechanismCatalytic Machinery Nuclear pre- mRNA Very common; used for most eukaryotic genes Two transesterificat ion reactions; branch site A Major spliceoso me Group II introns Rare; some eu- Karyotic genes from organelles and prokaryotes Same as pre- mRNA RNA enzyme encoded by intron (ribozyme) Group I introns Rare; nuclear rRNA in some eukaryotics, organlle genes, and a few prokaryotic genes Two transesterific- ation reactions; exogenous G Same as group II introns
 Group I introns Rare; nuclear rRNA in some eukaryotics, organlle genes, and a few prokaryotic genes Two transesterific- ation reactions; exogenous G Same as group II introns")
39
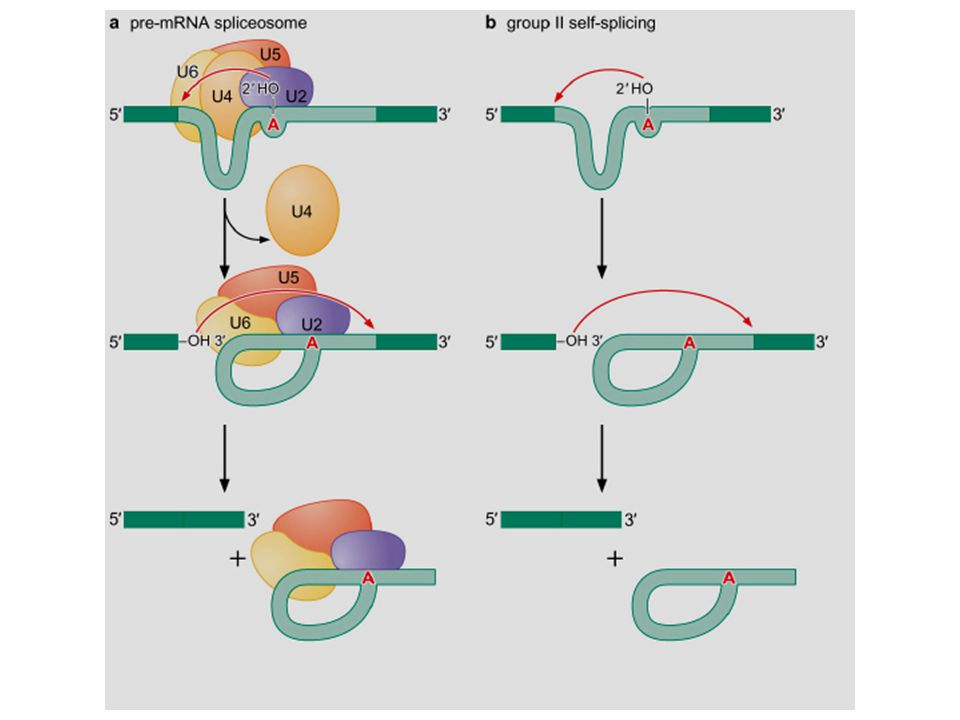
The chemistry of group II intron splicing and RNA intermediates produced are the same as that of the nuclear pre-mRNA.

41
Ⅲ Group I introns release a linear intron rather than a lariat

42
● Instead of using a branch point A, group I introns use a free G to attack the 5’ splice site. ● This G is attached to the 5’ end of the intron. The 3’-OH group of the 5’ exon attacks the 5’ splice site. ● The two-step transesterification reactions are the same as that of splicing of the group II intron and pre-mRNA introns.

43
G instead of A a linear introna Lariat intron

44
Group I introns 1. Smaller than group II introns. 2. Share a conserved secondary structure, which includes an “internal guide sequence” base-pairing with the 5’ splice site sequence in the upstream exon. 3. The tertiary structure contains a binding pocket that will accommodate the guanine nucleotide or nucleoside cofactor.

45
The similarity of the structures of group II introns and U2- U6 snRNA complex formed to process first transesterification

46
Ⅳ How does spliceosome find the splice sites reliably

47
● Two kinds of splice-site recognition errors Splice sites can be skipped. ● “Pseudo” splice sites could be mistakenly recognized, particularly the 3’ splice site.

49
Reasons for the recognition errors (1) The average exon is 150 nt, and the average intron is about 3,000 nt long (some introns are near 800,000 nt). It is quite challenging for the spliceosome to identify the exons within a vast ocean of the intronic sequences. (2) The splice site consensus sequence are rather loose. For example, only AG/G tri-nucleotides is required for the 3’ splice site, and this consensus sequence occurs every 64 nt theoretically.
 The splice site consensus sequence are rather loose. For example, only AG/G tri-nucleotides is required for the 3’ splice site, and this consensus sequence occurs every 64 nt theoretically..")
50
Two ways to enhance the accuracy of the splice- site selection 1. Because the C-terminal tail of the RNA polymerase II carries various splicing proteins, co- transcriptional loading of these proteins to the newly synthesized RNA ensures all the splice sites emerging from RNAP II are readily recognized, thus preventing exon skipping. 2. There is a mechanism to ensure that the splice sites close to exons are recognized preferentially. SR proteins bind to the ESEs (exonic splicing enhancers) present in the exons and promote the use of the nearby splice sites by recruiting the splicing machinery to those sites
 present in the exons and promote the use of the nearby splice sites by recruiting the splicing machinery to those sites.")
51
SR proteins, bound to exonic splicing enhancers (ESEs), interact with components of splicing machinery, recruiting them to the nearby splice sites.
, interact with components of splicing machinery, recruiting them to the nearby splice sites.")
52
SR proteins are essential for splicing 1. Ensure the accuracy and efficacy of constitutive splicing. 2. Regulate alternative splicing. 3. There are many varieties of SR proteins. Some are expressed preferentially in certain cell types and control splicing in cell-type specific patterns.

53
Topic Ⅳ Alternative Splicing

54
Ⅰ Single genes can produce multiple products by alternative splicing Many genes in higher eukaryotes encode RNAs that can be spliced in alternative ways to generate two or more different mRNAs and, thus, different protein products.

55
Drosophila DSCAM gene can be spliced in 38,000 alternative ways

56
There are five different ways to alternatively splice a pre-mRNA

57
Ⅱ Alternative splicing can be either constitutive or regulated

58
An example of constitutive alternative splicing : Splicing of the SV40 T antigen RNA

59
Alternative splicing is regulated by activators and repressors ● The regulating sequences : exonic (or intronic) splicing enhancers (ESE or ISE) or silencers (ESS and ISS). The former enhance and the latter repress splicing. ● Proteins that regulate splicing bind to these specific sites for their action.

60
SR proteins binding to enhancers act as activators. (1) One domain is the RNA-recognition motif (RRM). (2) The other domain is RS domain rich in arginine and serine. This domain mediates interactions between the SR proteins and proteins within the splicing machinery.
 One domain is the RNA-recognition motif (RRM). (2) The other domain is RS domain rich in arginine and serine. This domain mediates interactions between the SR proteins and proteins within the splicing machinery..")
61
hnRNPs binds RNA and act as repressors 1. Most silencers are recognized by hnRNP ( heterogeneous nuclear ribonucleoprotein) family. 2. These proteins bind RNA, but lack the RS domains. Therefore, (1) They cannot recruit the splicing machinery. (2) they block the use of the specific splice sites that they bind.
 family. 2. These proteins bind RNA, but lack the RS domains. Therefore, (1) They cannot recruit the splicing machinery. (2) they block the use of the specific splice sites that they bind..")
62
Regulated alternative splicing

63
Binds at each end of the exon and conceals it Coats the RNA and makes the exons invisible to the splicing machinery An example of repressors: inhibition of splicing by hnRNPI

64
The outcome of alternative splicing 1. Producing multiple protein products, called isoforms. 2. Switching on and off the expression of a given gene. In this case, one functional protein is produced by a splicing pattern, and the non-functional proteins are resulted from other splicing patterns.

65
Ⅲ A small group of intron are spliced by minor spliceosome ● This spliceosome works on a minority of exons, and those have distinct splice- site sequence. ● The chemical pathway is the same as the major spliceosome.

66
U11 and U12 are in places of U1 and U2, respectively

67
Topic Ⅴ Exon Shuffling

68
Exons are shuffled by recombin-ation to produce gene encoding new proteins All eukaryotes have introns, and yet these elements are rare in bacteria. Two likely explanations for these situation: 1. Introns early model – introns existed in all organisms but have been lost from bacteria. 2. Intron late model – introns never existed in bacteria but rather arose later in evolution.

69
Why have the introns been retained in eukaryotes?

70
1. The need to remove introns, allows for alternative splicing which can generate multiple proteins from a single gene. 2. Having the coding sequence of genes divided into several exons allows new genes to be created by reshuffling exon.

71
Three observations suggest exon shuffling actually occur: 1. The borders between exons and introns within a gene often coincide with the boundaries between domains within the protein encoded by that gene. 2. Many genes, and proteins they encode, have apparently arisen during evolution in part via exon duplication and divergence. 3. Related exons are sometimes found in unrelated genes.

72
For example: DNA-binding protein

73
Exons have been reused in genes encoding different proteins

74
Topic Ⅵ RNA Editing

75
RNA editing is another way of changing the sequence of an mRNA I. Site specific deamination 1. A specifically targeted C residue within mRNA is converted into U by the deaminase. 2. The process occurs only in certain tissues or cell types and in a regulated manner. 3. Adenosine deamination also occurs in cells. The enzyme ADAR (adenosine deaminase acting on RNA) convert A into Inosine. Insone can base-pair with C, and this change can alter the sequence of the protein. 4. An ion channel expressed in mammalian brains is the target of Adenosine deamination.
 convert A into Inosine. Insone can base-pair with C, and this change can alter the sequence of the protein. 4. An ion channel expressed in mammalian brains is the target of Adenosine deamination..")
77
The human apolipoprotein gene Stop code In liverIn intestines

78
II Guide RNA-directed uridine insertion or deletion 1. This form of RNA editing is found in the mitochondria of trypanosomes. 2. Multiple Us are inserted into specific region of mRNAs after transcription (or US may be deleted).
..")
79
3. The addition of Us to the message changes codons and reading frames, completely altering the “meaning” of the message. 4. Us are inserted into the message by guide RNAs (gRNAs).
..")
80
gRNAs Having three regions: anchor– directing the gRNAs to the region of mRNAs it will edit. editing region – determining where the Us will be inserted. poly-U stretch

82
Topic Ⅶ mRNA Transport

83
Once processed, mRNA is packaged and exported from the nucleus into the cytoplasm for translation All the fully processed mRNAs are transported to the cytoplasm for translation into proteins.

84
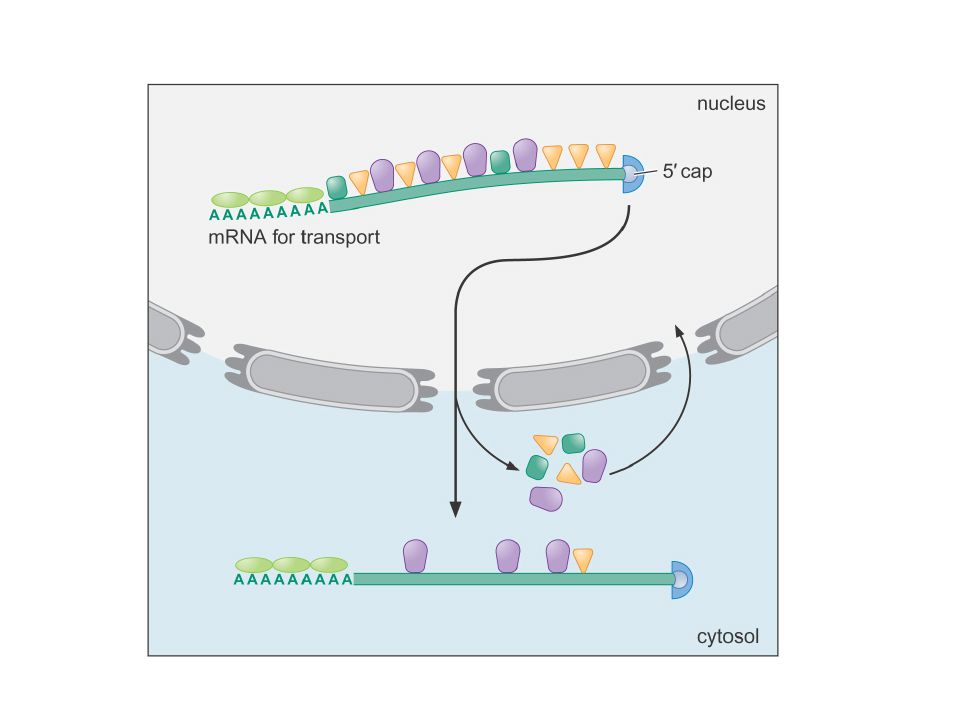
● Movement from the nucleus to the cytoplasm is an active and carefully regulated process. ● The damaged, misprocessed and liberated introns are retained in the nucleus and degraded. 1. A typical mature mRNA carries a collection of proteins that identifies it as being ready for transport. 2. Export takes place through the nuclear pore complex.

85
3. Once in the cytoplasm, some proteins are discarded and are then imported back to the nucleus for another cycle of mRNA transport. Some proteins stay on the mRNA to facilitate translation.

87
Summary 1. Why RNA splicing is important? 2. Chemical reaction: determination of the splice sites, the products, trans-splicing. 3. Spliceosome: splicing pathway and finding the splice sites. 4. Self-splicing introns and mechanisms. 5. Alternative splicing and regulation, alternative spliceosome. 6. Two different mechanisms of RNA editing. 7. mRNA transport-a link to translation.

Similar presentations








>")





>")


 MISS NUR SHALENA SOFIAN.>")


>")



 Umm-e-Habiba(BB307-035). Gene splicing “Gene splicing is the removal of introns from the primary trascript of a discontinuous gene.>")