Download presentation
Presentation is loading. Please wait.

1
Antibody Structure and the Generation of B-Cell Diversity
Chapter 2 Antibody Structure and the Generation of B-Cell Diversity

2
The Structural Basis of Antibody Diversity

3
Antibodies and Antigens
Five chemically and physically distinct classes of antibodies (IgG, IgA, IgM, IgD, IgE). Bind with high specificity and affinity. Produced by the B lymphocytes in response to infection. Circulate as a major component of the plasma in blood and lymph. Bind to pathogenic microorganism and their toxins in the extracellular spaces of the body. Antibody repertoire might be as high as 1016. Antigens…. Are bound by antibodies. Are biological macromolecules. proteins and carbohydrates most common antigens
. Bind with high specificity and affinity. Produced by the B lymphocytes in response to infection. Circulate as a major component of the plasma in blood and lymph. Bind to pathogenic microorganism and their toxins in the extracellular spaces of the body. Antibody repertoire might be as high as Antigens…. Are bound by antibodies. Are biological macromolecules. proteins and carbohydrates most common antigens.")
4
Antibody (Ab) - Immunglobulins
Antibodies are the secreted form of proteins known more generally as immunoglobulins (Igs) Mature B cells express Igs (B cell receptor) When Ag binds Igs, the B cell proliferates and then differentiates into a plasma cell Plasma cell secretes Ab with same specificity as the membrane-bound Igs
 Mature B cells express Igs (B cell receptor) When Ag binds Igs, the B cell proliferates and then differentiates into a plasma cell. Plasma cell secretes Ab with same specificity as the membrane-bound Igs.")
5
Antibody Functions Antibody functions in host defense:
Antibody recognizes and binds its corresponding antigen Antibody targets the bound antigen to other components of the immune system each of these functions is performed by different parts of the antibody molecule

6
Antibody Structure Antibodies are glycoproteins
Basic unit of four polypeptide chains (2 pairs L & H chains) covalent disulfide bridges noncovalent interactions Total MW~150kDa Two identical heavy chains (H chains) (50 kDa each) Two identical, smaller, light chains (L chains) (25 kDa each) Arms of Y L chains paired with N-terminal pair of H chain - Covalently linked by a disulfide bond. Stem of the Y consists of the paired C-terminal portions of the two-H chains
 covalent disulfide bridges. noncovalent interactions. Total MW~150kDa. Two identical heavy chains (H chains) (50 kDa each) Two identical, smaller, light chains (L chains) (25 kDa each) Arms of Y. L chains paired with N-terminal pair of H chain - Covalently linked by a disulfide bond. Stem of the Y. consists of the paired C-terminal portions of the two-H chains.")
7
Antibody Variability H and L chains are divided into V and C regions
The V region variability = great diversity concentrated in the N-terminal region paired V regions of a H and L chain form the antigen binding site two identical antigen binding sites The C region constant region determines the fate of the antigen less variation in amino acid sequence between antibodies N-terminal C-terminal

8
Antibody Affinity Affinity is the tightness of binding of an antibody binding site to an antigen. The tighter the binding, the less likely the antibody is to dissociate from antigen. Different antibodies to the same antigen vary considerably in their affinity for an epitope. Antibodies produced by a memory response have higher affinity than those in a primary response.

9
Antibody Valence and Avidity
Valence of an antibody is the maximum number of antigenic determinants with which an antibody can react. Multiple binding sites on an antibody dramatically increases its binding (avidity) to antigens on particles such as bacteria or virus. Example: two binding sites on IgG are 100 times more effective at neutralizing virus than two unlinked binding sites. Avidity is the firmness of association between a multideterminant antigen and the antibodies produced against it. This combined effect (avidity) results from synergy of the binding strengths of each binding site.
 to antigens on particles such as bacteria or virus. Example: two binding sites on IgG are 100 times more effective at neutralizing virus than two unlinked binding sites. Avidity is the firmness of association between a multideterminant antigen and the antibodies produced against it. This combined effect (avidity) results from synergy of the binding strengths of each binding site.")
10
Antibody Valence and Avidity

11
Proteases Cleave Antibody into Fab and Fc Fragments
Papain cleaves the Ig molecule into three pieces: Two Fab (Fragment Antigen Binding) fragments Antigen binding part One Fc fragment (Fragment Crystallizable) Part responsible for effector functions. Pepsin cleaves IgG to yield 1 F(ab’)2 fragment and the Fc fragment Fc fragment is broken into a number of smaller pieces
 fragments. Antigen binding part. One Fc fragment (Fragment Crystallizable) Part responsible for effector functions. Pepsin cleaves IgG to yield 1 F(ab’)2 fragment and the Fc fragment. Fc fragment is broken into a number of smaller pieces.")
12
Antibody Classes Different antibody classes with different biological activities have evolved to deal with diverse antigens. Heavy-chain constant regions define five main isotypes. Heavy chains , , , , IgA, IgD, IgE, IgG and IgM Each isotype has a different effector functions – with some overlap. Example: IgA is the most common antibody in mucosal secretions while IgM is mainly found in the plasma, and both are most effective at those locations

13
Isotypes of Heavy and Light Chains
Heavy-chain constant regions define five main isotypes of antibodies: IgA, IgD, IgE, IgG, and IgM Differences in length and sugars of heavy chain Only two isotypes of light chain kappa () and lambda () Each antibody has either or chains The antigen binding site is composed of two variable domains (VH and VL) Constant domain of light chain is CL Constant domain of the heavy chain is CH The heavy chain of IgG has three domains - CH1, CH2, CH3 Some other isotypes have four C domains
 and lambda () Each antibody has either or chains. The antigen binding site is composed of two variable domains (VH and VL) Constant domain of light chain is CL. Constant domain of the heavy chain is CH. The heavy chain of IgG has three domains - CH1, CH2, CH3. Some other isotypes have four C domains.")
14
IgG (MW~150 kD) Four different subclasses (IgG1, IgG2, IgG3 and IgG4)
Each subclass has slightly different H-chains and corresponding differences in their function. Found both in vascular and extravascular spaces as well as in secretions. Most abundant Ig in the blood. Provide the bulk of immunity to most blood borne infectious agents. The only antibody class to cross the placenta to provide passive humoral (innate) immunity to the developing and newborn infant.
 immunity to the developing and newborn infant.")
15
IgA (In Serum~170 kD, External Secretions ~420 kD Dimer)
Major Ig present in external secretions - colostrum, milk and saliva. In addition to the L-chains and the IgA heavy chain () secreted IgA also contains two other polypeptide chains- secretory component (SC) and J-chain (joining chain) SC is part of the molecule (poly-Ig receptor) involved in the transepithelial transport of exocrine IgA and stabilizes IgA against proteolytic degradation. The two four-chain units composing secretory IgA are held together by the J-chain through disulfide bridges. A first line of defense against microbes entering through mucosal surfaces. Respiratory, gastrointestinal and genitourinary tracts Secretory (dimeric) IgA is synthesized locally by plasma cells in mammary and salivary glands and along the respiratory, gastrointestinal and genitourinary tracts. IgA prevents colonization of mucosal surfaces by pathogens and mediates their phagocytosis.
 secreted IgA also contains two other polypeptide chains- secretory component (SC) and J-chain (joining chain) SC is part of the molecule (poly-Ig receptor) involved in the transepithelial transport of exocrine IgA and stabilizes IgA against proteolytic degradation. The two four-chain units composing secretory IgA are held together by the J-chain through disulfide bridges. A first line of defense against microbes entering through mucosal surfaces. Respiratory, gastrointestinal and genitourinary tracts. Secretory (dimeric) IgA is synthesized locally by plasma cells in mammary and salivary glands and along the respiratory, gastrointestinal and genitourinary tracts. IgA prevents colonization of mucosal surfaces by pathogens and mediates their phagocytosis.")
16
(1) Protected from proteases (5) (2) (3) (4) Receptor-mediated endocytosis
 Protected from proteases (5) (2) (3) (4) Receptor-mediated endocytosis")
17
IgM (~900 kD) IgM is the first antibody produced by B cells.
Is the antigen receptor (B cell receptor). Expressed as a four-chain unit - two H-chains and two L-chains. Also present as a soluble molecule in the blood. J-chain is associated with IgM in the blood and initiates the polymerization of its subunits. In the circulation, IgM is composed of five four-chain units with 10 combining sites (recall avidity slide). Has high avidity for antigens and is very efficient per molecule in dealing with pathogens especially early in the immune response before sufficient quantities of IgG have been produced.
. Expressed as a four-chain unit - two H-chains and two L-chains. Also present as a soluble molecule in the blood. J-chain is associated with IgM in the blood and initiates the polymerization of its subunits. In the circulation, IgM is composed of five four-chain units with 10 combining sites (recall avidity slide). Has high avidity for antigens and is very efficient per molecule in dealing with pathogens especially early in the immune response before sufficient quantities of IgG have been produced.")
18
IgD Present in low quantities in the circulation
(0.3 mg/ml in adult serum) IgD functions primarily as an antigen receptor on B cells. B cells express both IgM and IgD that are specific for the same antigen. When IgM and IgD on a B cell interact with an antigen, the antigen is internalized and processed and presented to helper T cells. This triggers B cells to proliferate and differentiate into plasma cells, thus initiating the development of a humoral immune response Humoral-pertains to extracellular fluid including the plasma and lymph.
 IgD functions primarily as an antigen receptor on B cells. B cells express both IgM and IgD that are specific for the same antigen. When IgM and IgD on a B cell interact with an antigen, the antigen is internalized and processed and presented to helper T cells. This triggers B cells to proliferate and differentiate into plasma cells, thus initiating the development of a humoral immune response. Humoral-pertains to extracellular fluid including the plasma and lymph.")
19
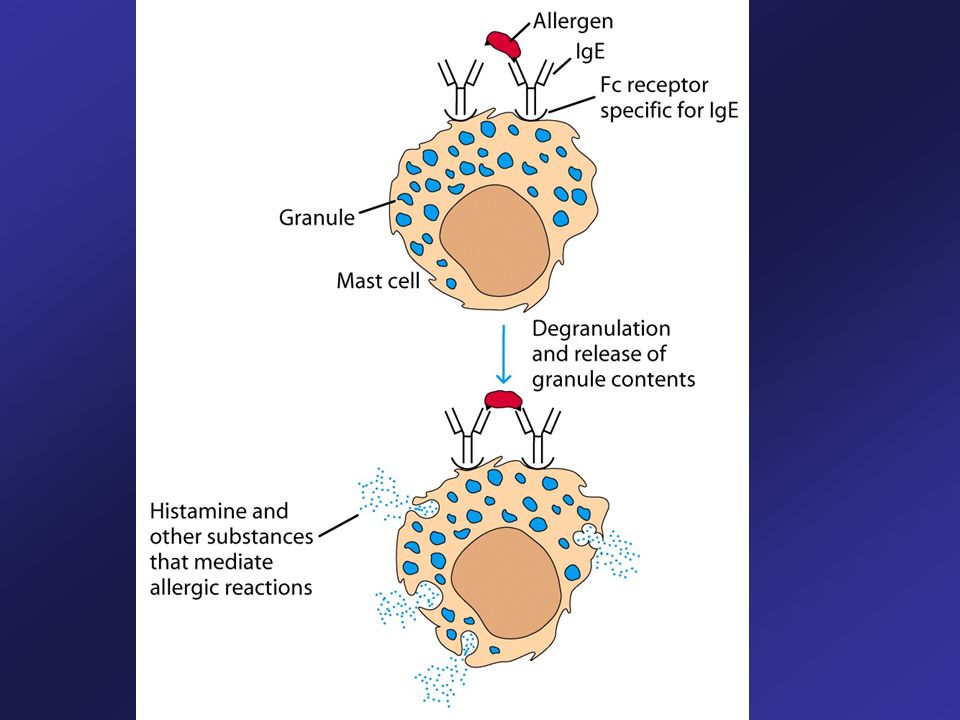
IgE Present in serum at very low levels nanograms per milliliter
Plays significant roles in enhancing acute inflammation protection from infection by worms Allergic reactions are predominantly associated with IgE After antigen stimulation induces plasma cells to produce IgE IgE binds to receptors on mast cells which are specific for the Fc regions of IgE. Antigen reintroduced into a previously sensitized individual binds to IgE on ‘armed’ mast cells and triggers release of the pharmacologically active agents – histamine (involved in immediate hypersensitivity such as hay fever and asthma).
.")
21
Structural Organization of Human Immunoglobulin Isotypes
Differences in length of the heavy-chain C regions and locations of the disulfide bonds. The hinge region present in IgG, IgA, IgD but not in IgM, IgE. Isotypes differ in the distribution of N-linked carbohydrate groups All occur as monomers in their membrane-bound form. Soluble, secreted form of IgD, IgE and IgG are always monomers. IgA forms monomers and dimers. IgM forms pentamers.

22
Immunoglobulins are flexible
The hinge region is a flexible tether, allowing independent movement of the two Fab arms. The angle between the arms can vary from 0° in dimers, to 60° or 90° degrees in triangles or squares.

23
Immunoglobulin Chains are Folded into Compact and Stable protein Domains
The V region at the N-terminal end of each H or L chain is composed of a single variable domain (V domain, VH or VL). VH and VL domains together form an antigen-binding site. The other domains have little or no sequence diversity within a particular isotype and are termed the constant domains (C domains) . Constant region of a light chain is composed of a single CL domain Constant region of a H chain is composed of three or four C domains (isotype dependent) the heavy chains of IgG have 3 domains-Ch1, CH2, Ch3 other isotypes have four C domains (previous slide) In the complete IgG molecule the pairing of the four polypeptide chains produces three globular regions, corresponding to the two Fab arms and the Fc stem (refer to fig 2.5)
. VH and VL domains together form an antigen-binding site. The other domains have little or no sequence diversity within a particular isotype and are termed the constant domains (C domains) . Constant region of a light chain is composed of a single CL domain. Constant region of a H chain is composed of three or four C domains (isotype dependent) the heavy chains of IgG have 3 domains-Ch1, CH2, Ch3. other isotypes have four C domains (previous slide) In the complete IgG molecule the pairing of the four polypeptide chains produces three globular regions, corresponding to the two Fab arms and the Fc stem (refer to fig 2.5)")
25
Immunoglobulin Chains are Folded into Compact and Stable protein Domains
Immunoglobulin-like domain = Immunoglobulin superfamily

26
Ag-binding Site is Formed from the Hypervariable Regions of a Heavy Chain and a Light Chain V Domain
V domains of light and heavy chains have hypervariable regions (HV), flanked by framework regions. Three hypervariable regions in each domain.
, flanked by framework regions. Three hypervariable regions in each domain.")
27
The Hypervariable Regions of Antibody V Domains Lie in Discrete Loops at One End of the Domain Structure The hypervariable regions of antibody V domains lie in discrete loops at the end of the domain. The pairing of the heavy and light chains bring together the hypervariable loops and create a composite hypervariable surface that forms the antigen-binding site. This surface determines specificity and diversity. The hypervariable loops are also called the complementarity-determining regions (CDRs).
.")
28
Epitopes for Antibody Binding are Exposed
Antibodies that are most effective are those that bind the exposed surface of a pathogen. Epitope is the part of the antigen to which an antibody binds. An antigen that contains more than one epitope is called multivalent. Poliovirus

29
Physical Properties of Antibody Binding Sites
The antibody binds to an antigenic determinant or epitope, commonly: glycoproteins, polysaccharides, glycolipids, and proteoglycans. Epitopes can be linear, or discontinuous (when different parts of the antigen are folded together). The binding forces are non-covalent: electrostatic, hydrogen bonds, van der Waals forces and hydrophobic forces. The binding strength is known as the affinity of the antibody for the antigen.
. The binding forces are. non-covalent: electrostatic, hydrogen bonds, van der. Waals forces and. hydrophobic forces. The binding strength is known as the affinity of the antibody for the antigen.")
30
Epitopes Can Bind to Pockets, Grooves or Extended Surfaces in the Antibody Binding Sites
The type of antigen bound by an antibody depends on the shape of the antigen-binding site. Antigens that are small molecules can be bound within deep pocket. Linear epitopes from proteins or carbohydrates can be bound within clefts or grooves. Binding of conformation epitopes of folded proteins takes place over an extended surface area.

31
Antibodies Binding to Three different Types of Epitope
Peptide epitope (red) bound to pocket in CDR loops Peptide epitope in groove between two V domains Fab binding to surface epitope of lysozyme
 bound to pocket in CDR loops. Peptide epitope in groove between two V domains. Fab binding to surface epitope of lysozyme.")
32
Monoclonal Antibodies
1975 Kohler and Milstein developed a procedure to create cell lines producing predetermined, monospecific and monoclonal antibodies (mAb). Standardized procedures involving fusion of an immortal cell (i.e. myeloma tumor cell) with a specific predetermined antibody-producing B cell from immunized animals or humans to create hybridoma cells producing monospecific and monoclonal antibodies (mAb). Monoclonal antibodies important in a variety of therapeutic, diagnostic and research settings.
. Standardized procedures involving fusion of an immortal cell (i.e. myeloma tumor cell) with a specific predetermined antibody-producing B cell from immunized animals or humans to create hybridoma cells producing monospecific and monoclonal antibodies (mAb). Monoclonal antibodies important in a variety of therapeutic, diagnostic and research settings.")
33
Production of Monoclonal Antibodies
Produced from a single clone from antibody-producing cells and thus have identical Ag-binding sites and identical isotypes. Antibodies can be humanized (mouse CDRs into human antibody) for therapeutic purposes.
 for therapeutic purposes.")
34
The flow cytometer allows individual cells to be identified by surface molecules.

35
Generation of Immunoglobulin Diversity

36
In all cells (except B cells) Ig genes are in a fragmented form that cannot be expressed.
The Ig H-chain and L-chain loci consists of families of gene segments containing alternative versions of parts of the Ig variable region. Germ-line DNA contains multiple gene segments encoding portions of a single Ig heavy or light chain. Germ-line DNA Rearranged Functional DNA

37
In B cell germ-line DNA gene segments are rearranged into functional genes.
Mature B cells contain chromosomal DNA that is no longer identical to germ-line DNA. Genomic rearrangement is an essential feature of lymphocyte differentiation. Germ-line DNA Rearrangement Rearranged Functional DNA

38
Generation of Diversity: Antibody Genes
DNA encoding Igs is found in three unlinked gene groups. One group encodes kappa () L-chains One lambda () L-chains One H-chains Each L-chain gene groups has multiple different copies of V gene segments and J segments. In addition, in the kappa () chain group there is one gene segment encoding the constant regions of kappa () chains. In the lambda () groups there are four lambda () chain constant regions gene segments. The H-chain gene groups has multiple different copies of V, D, and J gene segments and one gene segment for each of the constant regions for the different antibody classes and subclasses.
 L-chains. One lambda () L-chains. One H-chains. Each L-chain gene groups has multiple different copies of V gene segments and J segments. In addition, in the kappa () chain group there is one gene segment encoding the constant regions of kappa () chains. In the lambda () groups there are four lambda () chain constant regions gene segments. The H-chain gene groups has multiple different copies of V, D, and J gene segments and one gene segment for each of the constant regions for the different antibody classes and subclasses.")
40
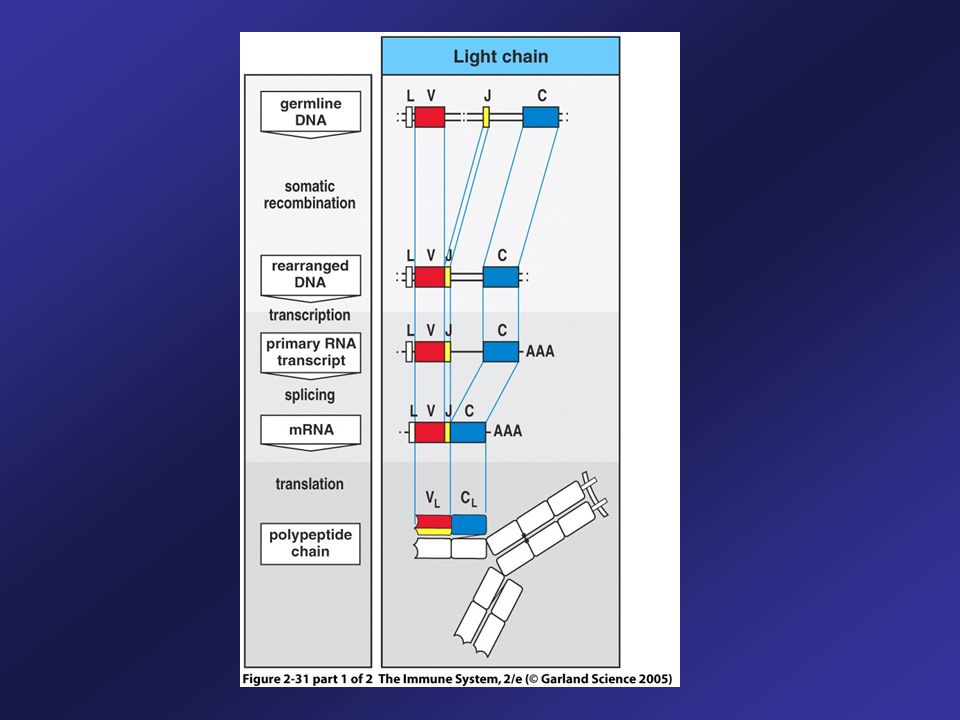
Generation of Diversity: Gene Rearrangement
During development, a single B cell randomly selects from its H-chain gene group, one V, one D and one J gene segment for rearrangement. The B cell then selects a L-chain from the kappa () or lambda () gene group, selecting one V and one gene J gene segment for rearrangement. These gene segments then recombine to create a gene (VJ) encoding the L chain variable region and a gene (VDJ) encoding the H chain variable region.
 or lambda () gene group, selecting one V and one gene J gene segment for rearrangement. These gene segments then recombine to create a gene (VJ) encoding the L chain variable region and a gene (VDJ) encoding the H chain variable region.")
41
Generation of Ig Diversity in B Cells Prior to Contact with Antigen
In the germ line configuration, the Ig genes are fragmented into gene segments along the chromosome. Individual gene segments must be rearranged. This only occurs in B cells. It occurs during their development from precursors in the bone marrow. When gene rearrangements complete, heavy and light chains are produced and membrane bound Ig is expressed on the cell surface. The B cell can now recognize and respond to Ag.

44
Chromosomal Arrangement of Ig Genes
Heavy-chain locus on chromosome 14; light-chain on chromosome 2; light-chain on chromosome 22. Different segments encode leader peptide (L), variable region (V) and constant regions (C). Heavy-chain locus contains C genes for all isotypes. The C genes are ready to be transcribed. The V genes first undergo rearrangement.
, variable region (V) and constant regions (C). Heavy-chain locus contains C genes for all isotypes. The C genes are ready to be transcribed. The V genes first undergo rearrangement.")
45
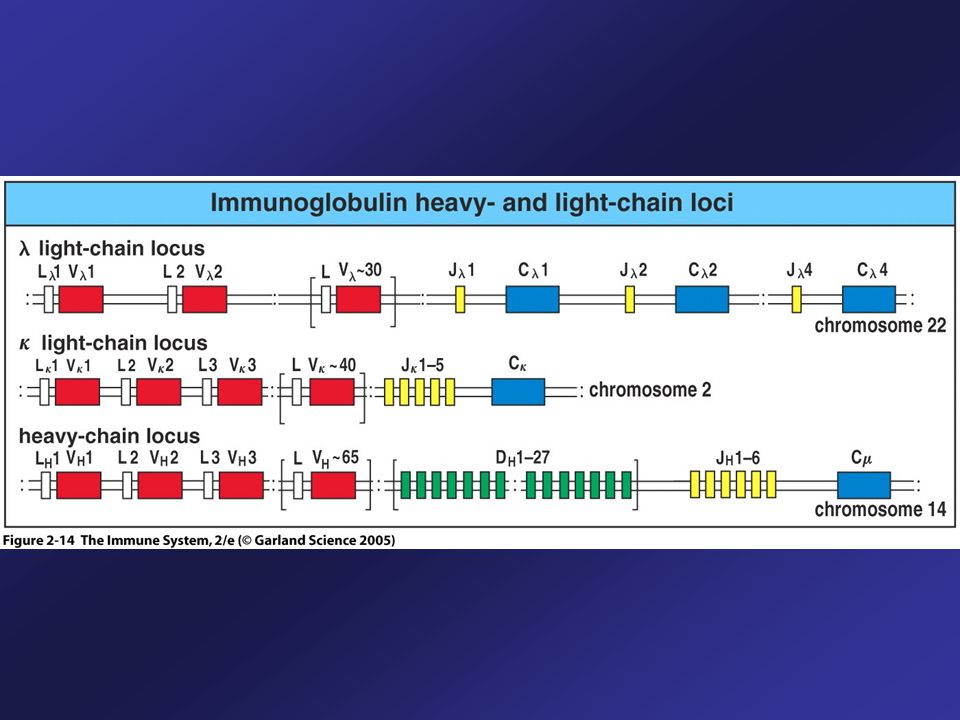
Immunoglobulin Heavy- and Light–Chain Loci
The V regions of light-chain comprised of two segments: variable (V) and joining (J). Heavy chain also contains D segments between V and J. First and second hypervariable region of light-chain (CDR1, 2) are encoded by different V segments, the third CDR is determined by the V/J junction.
 and joining (J). Heavy chain also contains D segments between V and J. First and second hypervariable region of light-chain (CDR1, 2) are encoded by different V segments, the third CDR is determined by the V/J junction.")
46
The Germline Organization of the Human Immunoglobulin Heavy-Chain and Light-Chain Loci

47
Somatic Recombination of Ig
All cells V-region sequences are constructed from gene segments L-chain, single recombination VJ H-chain, 2 recombinations: first DJ, then VDJ. C-regions at not rearranged at this point. B cells

48
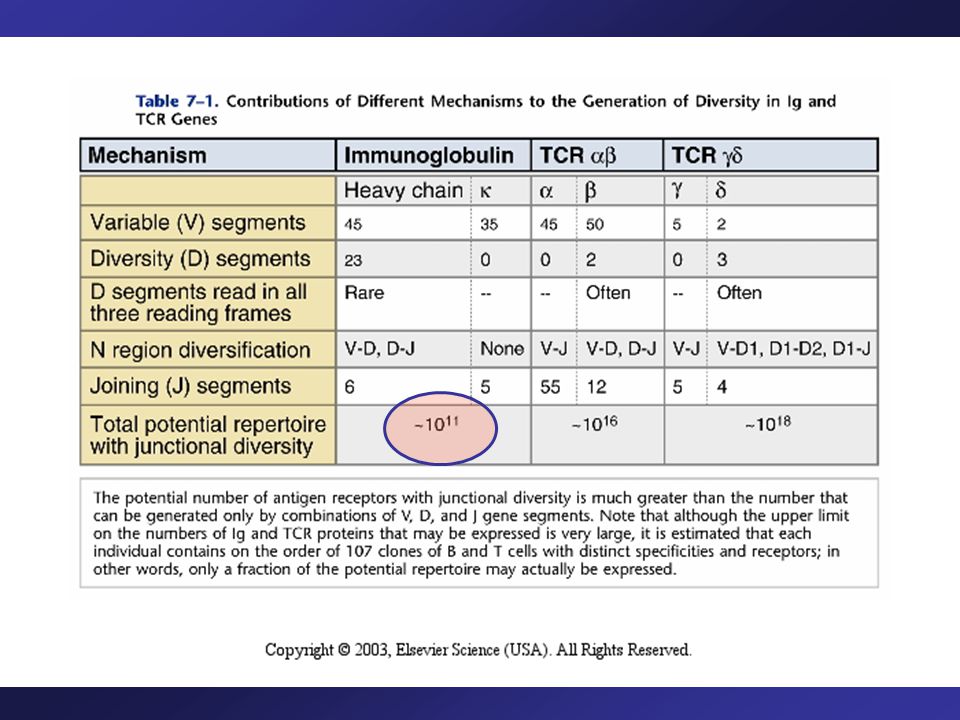
#’s of Functional Gene Segments Used to Construct the Variable Regions of Human Ig H and L Chains
The V, J (and D) segments are combined at random, thus many variations possible. Human light chain: 40 V segments and 5 J segments - therefore 200 combinations (40 x 5). Human light-chain: 30 x 4 = 120. Human heavy-chain: 65 V, 27 D, 6 J (65 x 27 x 6 = 10,530 combinations).
 segments are combined at random, thus many variations possible. Human light chain: 40 V segments and 5 J segments - therefore 200 combinations (40 x 5). Human light-chain: 30 x 4 = 120. Human heavy-chain: 65 V, 27 D, 6 J (65 x 27 x 6 = 10,530 combinations).")
50
Mechanism of V-region DNA Rearrangements
There are unique recombination signal sequences (RSSs) flanking each germ-line V, D, and J gene segment. One RSS is located 3’ to each V gene segment and 5’ to each J gene segment, and on both sides of each D gene segment. These sequence are signals for the recombination process that rearranges the gene.
 flanking each germ-line V, D, and J gene segment. One RSS is located 3’ to each V gene segment and 5’ to each J gene segment, and on both sides of each D gene segment. These sequence are signals for the recombination process that rearranges the gene.")
51
Each RSS contains conserved palinodromic heptamer and a conserved AT-rich nonamer sequence, separated by an intervening sequence of 12 or 23 base pairs. The intervening base pairs correspond to one or two turns of the DNA helix. Two turns of DNA helix One turn of DNA helix

52
Each V, D, Or J Gene Segment is Flanked by Recombination Signal Sequences
Signal sequences having 1-turn spacer can only join with sequences having 2-turn spacers – 1-turn/2-turn joining rule. 2-turn RSS 1-turn RSS 1-turn RSS 2-turn RSS 2-turn RSS 1-turn RSS 1-turn RSS 2-turn RSS

53
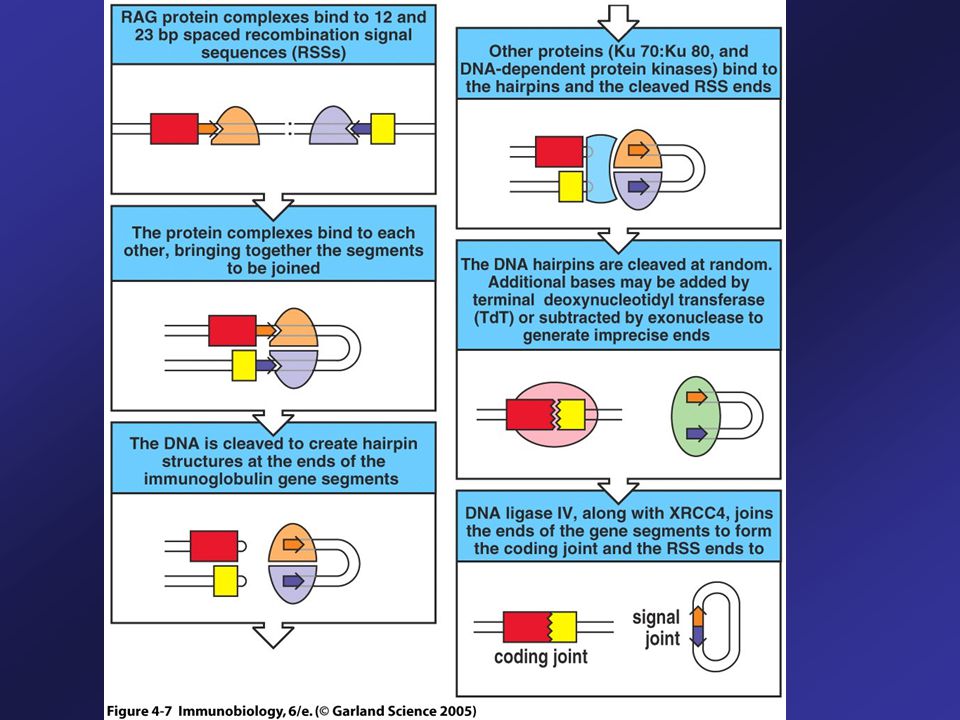
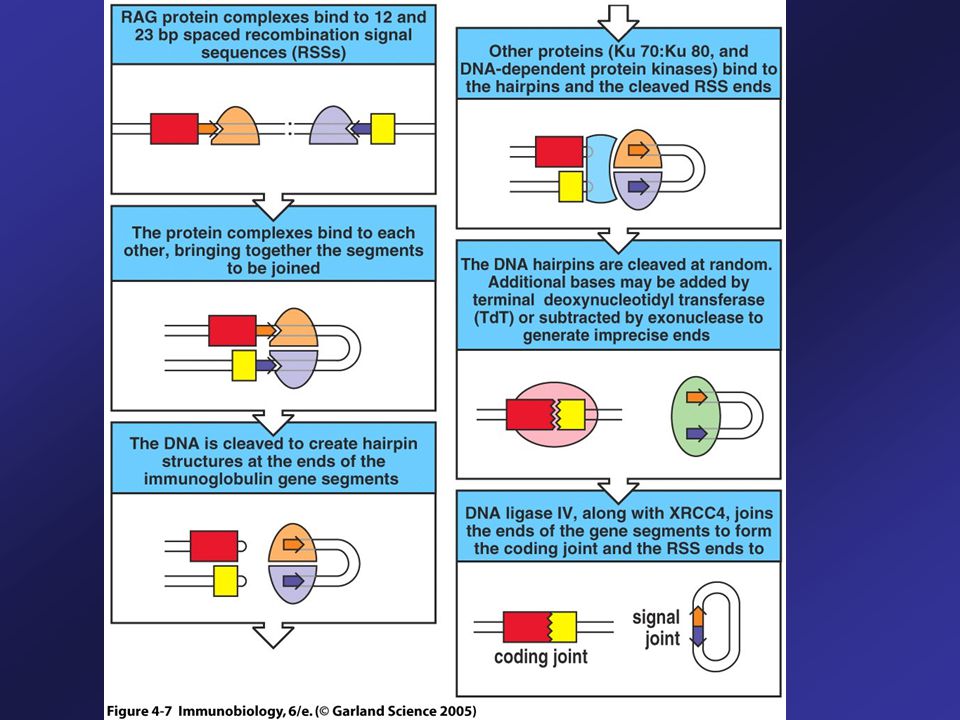
Recombination Enzymes
Set of enzymes needed to recombine V, D, and J gene segments is called V(D)J recombinase. Two components of the recombinase – RAG-1 and RAG-2 are only made in lymphocytes. Other enzymes required are present in all cells: DNA ligase IV, DNA-dependent protein kinase (DNA-PK) and Ku protein. First step in gene rearrangement is the binding of the RAG complex to the RSS.
J recombinase. Two components of the recombinase – RAG-1 and RAG-2 are only made in lymphocytes. Other enzymes required are present in all cells: DNA ligase IV, DNA-dependent protein kinase (DNA-PK) and Ku protein. First step in gene rearrangement is the binding of the RAG complex to the RSS.")
54
Gene Segments Encoding the Variable Region are Joined by Recombination at Recombination Signal Sequences A RAG complex binds to the 23-bp spacer (2-turn RSS) and another to the 12-bp spacer (1-turn-RSS), so that a RSS containing a 12-bp spacer is brought together with that containing a 23-bp spacer.
 and another to the 12-bp spacer (1-turn-RSS), so that a RSS containing a 12-bp spacer is brought together with that containing a 23-bp spacer.")
55
1-turn/2-turn joining Rule (12/23 Rule)
12/23 rule ensures that gene segments are joined in the correct order. The DNA molecules are broken at the ends of the heptamer sequences (orange) and are then joined together.
 and are then joined together.")
56
Different Topologies, Signal and Coding Joints
The DNA molecules are broken at the ends of the heptamer sequences (orange) and are then joined together with differ topologies. The region of DNA that was originally between the V and J segments to be joined is excised as a small circle of DNA that has no function (Signal joint). Within the chromosomal DNA, the V and J segments are joined to form the coding joint.
 and are then joined together with differ topologies. The region of DNA that was originally between the V and J segments to be joined is excised as a small circle of DNA that has no function (Signal joint). Within the chromosomal DNA, the V and J segments are joined to form the coding joint.")
58
Recombination Enzymes Produce Additional Diversity in Ag-binding Sites
The enzymes that cut and splice DNA during recombination introduce additional nucleotides not encoded in germ line DNA. Known as junctional diversity. Important source of Ig variability - adds a factor of 3 x 107 to overall diversity.

59
If one allele rearranges non-productively, then the B cell rearranges the other allele.
If this also fails and no rearranged heavy or light chain results the cell will die.

60
B cells are diploid and contain both maternal and paternal chromosomes
B cells are diploid and contain both maternal and paternal chromosomes. Rearranged heavy chain or light is expressed from only one chromosome. This is called allelic exclusion.

61
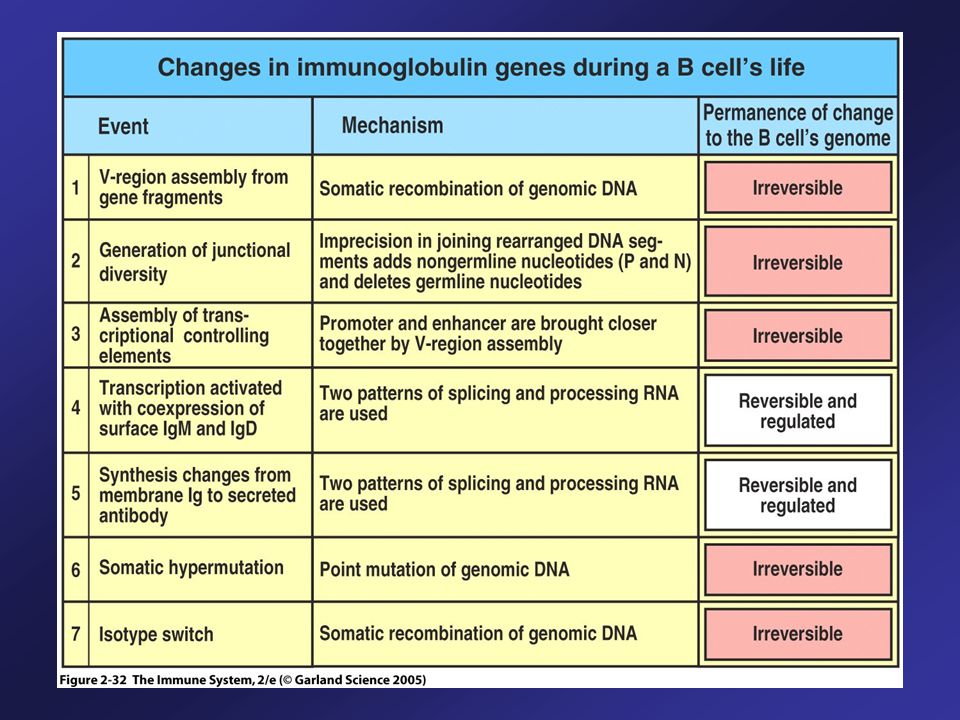
Diversity of Immunoglobulin is Generated by Multiple Processes
Multiple germ-line gene segments Combinatorial V-(D)-J joining Junctional diversity P-region nucleotide addition N-region nucleotide addition Somatic hypermutation Combinatorial association of heavy and light chains Subsequent joining of coding sequences is imprecise.
-J joining. Junctional diversity. P-region nucleotide addition. N-region nucleotide addition. Somatic hypermutation. Combinatorial association of heavy and light chains. Subsequent joining of coding sequences is imprecise.")
63
Mature B cells produces
both IgM and IgD with identical antigen specificity.

64
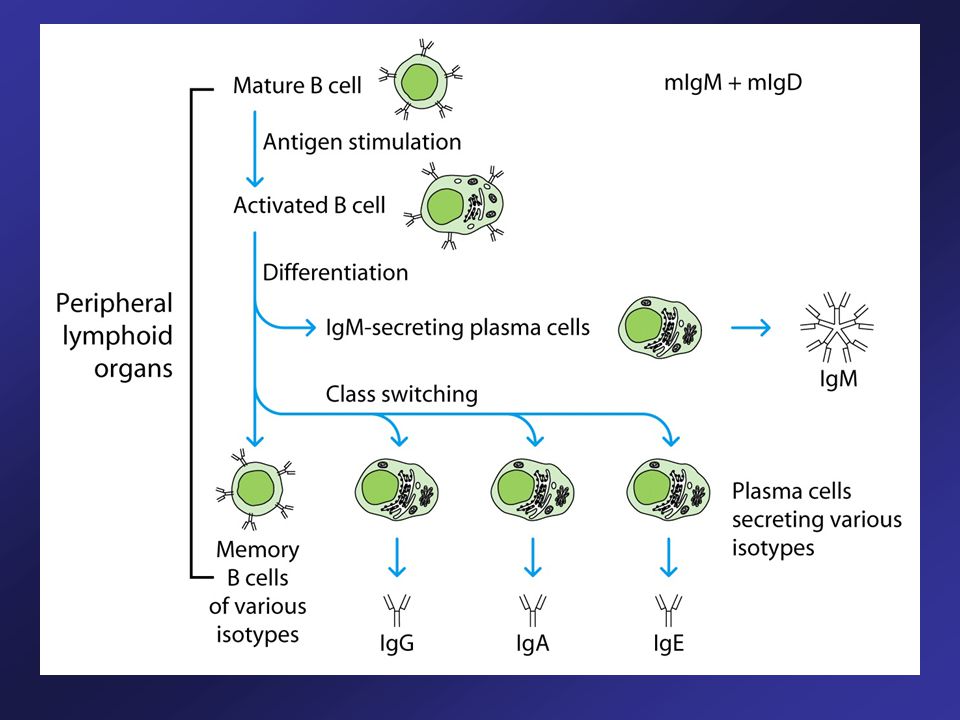
Naïve B Cells Use Alternative Splicing to Make Both IgM and IgD
Rearrangement of V, D, and J segments produces a functional heavy chain gene. The same assembled V-region serves all the heavy-chain C genes. The B cell first expresses IgM and IgD ( and nearest to assembled VDJ).
.")
65
Expression of IgM and IgD
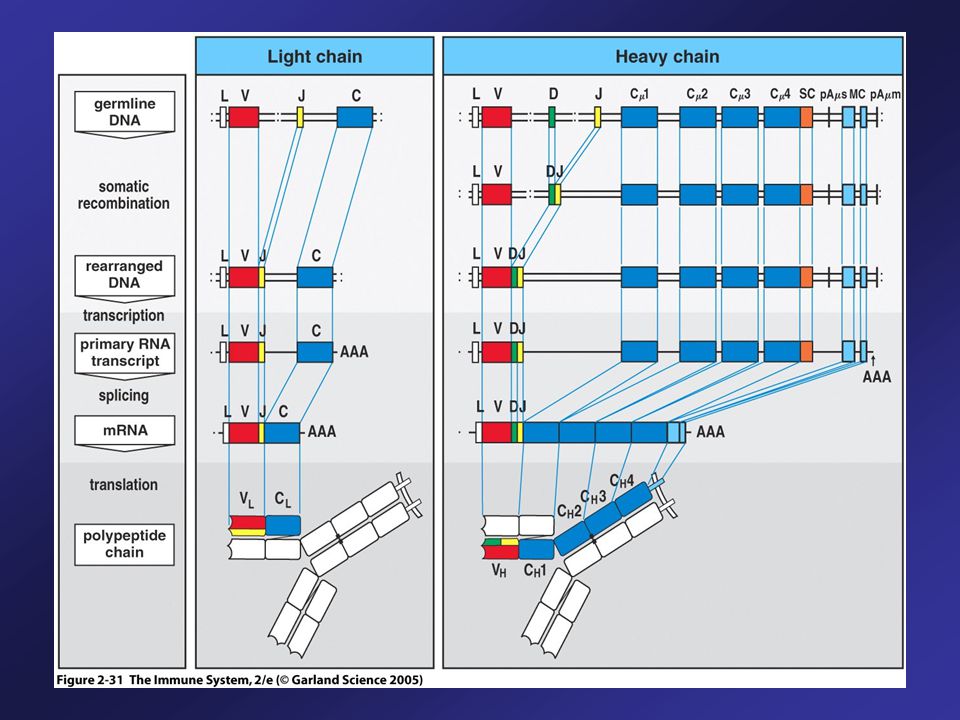
Transcription begins with VH promoter and extends through and C genes. This long primary transcript is processed by cleavage, polyadenylation and splicing. Cleavage and polyadenylation of the site (pA) and splicing between C exons yields mRNA encoding heavy chain for IgM. Cleavage and polyadenylation of the site (pA) and splicing between C exons yields mRNA encoding heavy chain for IgD. Upon encountering a specific antigen, many B cells change the isotype that they produce and produce IgG, or IgA, or IgE.
 and splicing between C exons yields mRNA encoding heavy chain for IgM. Cleavage and polyadenylation of the site (pA) and splicing between C exons yields mRNA encoding heavy chain for IgD. Upon encountering a specific antigen, many B cells change the isotype that they produce and produce IgG, or IgA, or IgE.")
66
Expression of IgM and IgD

67
The Functional B Cell Receptor
Membrane-bound Igs are associated with two other proteins: Ig and Ig Ig and Ig have longer cytoplasmic tails than Ig and interact with intracellular signaling molecules

68
Diversification of Antibodies After B Cells Encounter Ag
A B cell’s first encounter with antigen triggers proliferation, differentiation and secretion of antibodies.

69
Production of Secreted Antibodies
Gene rearrangement in immature B cell leads to expression of functional heavy and light chains. Mature (naïve) B cell expresses surface IgM and IgD. After encountering Ag, IgM, and IgD produced as secreted Abs. IgM important; IgD has unknown function. All classes of Ig can be made as membrane form and secreted form (the membrane binding domain edited out by alternative splicing). Mature plasma cell only produces secreted antibody.
 B cell expresses surface IgM and IgD. After encountering Ag, IgM, and IgD produced as secreted Abs. IgM important; IgD has unknown function. All classes of Ig can be made as membrane form and secreted form (the membrane binding domain edited out by alternative splicing). Mature plasma cell only produces secreted antibody.")
70
Production of Secreted Antibodies
Difference between membrane-bound and secreted antibody lies in the Carboxy-terminus of H-chain. Secreted antibody has a hydrophilic sequence.

71
Production of Membrane Antibodies
Membrane-associated antibody has a hydrophobic anchor sequence that is inserted into membranes. Difference is determined by different pattern of RNA processing.

72
Expression of the membrane or secreted forms of and heavy chains depends on polyadenylation sites. Secreted IgM Membrane IgM

73
Expression of the membrane or secreted forms of and heavy chains depends on polyadenylation sites. Secreted IgM Membrane IgD

74
Rearranged V-region Sequences Further Diversified by Somatic Hypermutation
Once B cell has been activated by Ag, further diversification of the V-domain coding region occurs. Called somatic hypermutation - introduces point mutations throughout rearranged V regions at very high rate – more than a million times greater than the ordinary mutation rate. Acts selectively at certain types of DNA sequences commonly found in CDRs. Mechanism involves activation-induced cytidine deaminase (AID) – an ezyme that converts cytidine to uracil, which are the excised and replaced with non-templated nucleotides.
 – an ezyme that converts cytidine to uracil, which are the excised and replaced with non-templated nucleotides.")
75
Somatic Hypermutation Introduces Diversity into Expressed Immunoglobulin Genes
Antibodies, raised by immunization with the same antigen, were collected 1 and 2 weeks after immunization and amino acid sequences were determined. Each red line represents one antibody and the red bars represent amino acid positions that differ from the prototypic sequence. One week after primary immunization most of the antibodies were IgM and showed very little sequence variation in the V region. Two weeks after immunization, both IgG and IgM were represent and all six CDRs were affected.

76
1 Week Versus 2 Weeks Post Primary Immunization

77
Affinity Maturation Thus somatic hypermutation results in mutant Igs on the surface of B cells. Some of these mutants will have a higher affinity for the Ag - they will therefore be more likely to bind the Ag, and B cells bearing these molecules will be preferentially selected to mature into Ab-secreting cells. As immune response proceeds, antibodies of higher affinity will be produced. Called “affinity maturation”

78
Isotype Switching Produces Igs with Different C Regions but Identical Ag Specificities
IgM is the first Ab produced in an immune response. Secreted as a circular pentamer; has 10 binding sites; therefore binds tightly to Ag; limited in effector mechanisms.

79
Switch Sequences or Switch Regions
Antibodies with other effector mechanisms are produced by isotype switching or class switching. Involves a further DNA recombination event that places the original V-region with other heaving C-regions. Switching is accomplished by recombination within a cluster of C genes, excising the previous C genes and joining a new C gene with previously assembled V-region. Antigen specificity thus remains the same but the isotype of the antibody changes. Flanking the 5’ side of each C gene, with the exception of the gene, are highly repetitive sequences that mediate recombination – called switch regions or sequences.

80
Switch Sequences or Switch Regions

81
The Physical Properties of the Human Immunoglobulin Isotypes
Antibodies with different constant regions have different effector functions. Humans have five classes of Ig: IgA, IgD, IgE, IgG, and IgM. Classes further divided into subclasses: IgG1, IgG2, IgG3, IgG4.

82
Antibodies Act on Pathogens in Various Ways
Neutralizing antibodies directly inactivates pathogen. For example, virus prevented from binding its receptor Opsonization - facilitate phagocytosis. IgM first antibody produced against pathogen - low affinity, therefore multiple binding sites needed for strong binding. Ag binding exposes sites on the C region that permit complement fixation. This results in opsonization or direct killing by complement.

83
IgM First antibody produced.
Principly made by plasma cells in lymphoid organs. On initial immune response most of the antibody will be of low affinity and the multi-binding sites on IgM are needed.

84
Somatic Hypermutation Leads to Antibodies with Increased Affinity
After affinity maturation, two Ag binding sites are sufficient to produce strong binding. By switching isotype, different effector functions are brought into play while preserving antigen specification.

85
IgG IgM gives way to IgG IgG most abundant antibody in the internal fluids. IgG more flexible. Also implement more effector functions. IgG bound by Fc receptors, can activate complement, can cross the placenta to provide fetus protective antibodies from mother.

86
There are four human IgG subclasses,
distinguished by the chain sequences.

87
IgA Monomeric IgA found in the circulation.
Dimeric IgA is made in lymphoid tissue underlying mucosal surfaces and is the antibody secreted into the lumen of the gut. The principle antibody in secretions, including milk, tears, saliva and sweat. Mucosa is a major area of intersection between pathogens and the immune system. More IgA is made than any other isotype.

88
IgE IgE binds to high affinity receptor on mast cells.
Ag binding provokes a strong inflammatory reaction. Involved in expulsion of worms and other parasites. Triggers allergic reactions.

89
Each Immunoglobulin Isotype has Specialized Functions and Distinct Properties

91
Dealing with Chapter 2 OVERVIEW

92
Proteases Cleave Antibody into Fab and Fc Fragments
Papain cleaves the Ig molecule into three pieces: Two Fab (Fragment Antigen Binding) fragments Antigen binding part One Fc fragment (Fragment Crystallizable) Part responsible for effector functions. Pepsin cleaves IgG to yield 1 F(ab’)2 fragment and the Fc fragment Fc fragment is broken into a number of smaller pieces
 fragments. Antigen binding part. One Fc fragment (Fragment Crystallizable) Part responsible for effector functions. Pepsin cleaves IgG to yield 1 F(ab’)2 fragment and the Fc fragment. Fc fragment is broken into a number of smaller pieces.")
93
Structural Organization of Human Immunoglobulin Isotypes
Differences in length of the heavy-chain C regions and locations of the disulfide bonds. The hinge region present in IgG, IgA, IgD but not in IgM, IgE. Isotypes differ in the distribution of N-linked carbohydrate groups All occur as monomers in their membrane-bound form. Soluble, secreted form of IgD, IgE and IgG are always monomers. IgA forms monomers and dimers. IgM forms pentamers.

94
Antibodies Binding to Three different Types of Epitope
Peptide epitope (red) bound to pocket in CDR loops Peptide epitope in groove between two V domains Fab binding to surface epitope of lysozyme
 bound to pocket in CDR loops. Peptide epitope in groove between two V domains. Fab binding to surface epitope of lysozyme.")
95
The Germline Organization of the Human Immunoglobulin Heavy-Chain and Light-Chain Loci

96
Somatic Recombination of Ig
V-region sequences are constructed from gene segments L-chain, single recombination VJ H-chain, 2 recombinations: first DJ, then VDJ

97
Each V, D, Or J Gene Segment is Flanked by Recombination Signal Sequences
Two types of RSS exist One consist of a nonamer (9bp) and a heptamer (7bp) separated by a spacer of 12bp The other consists of the same 9- and 7-bp separated by a 23-bp spacer
 and a heptamer (7bp) separated by a spacer of 12bp. The other consists of the same 9- and 7-bp separated by a 23-bp spacer.")
99
Recombination Enzymes Produce Additional Diversity in Ag-binding Sites
The enzymes that cut and splice DNA during recombination introduce additional nucleotides not encoded in germ line DNA Known as junctional diversity Important source of Ig variability - adds a factor of 3 x 107 to overall diversity

101
Expression of IgM and IgD
Transcription begins with VDJ and includes and C genes. This primary RNA transcript is spliced to produce IgM or IgD mRNA Naïve B cells express both IgM and IgD on their surfaces Upon encountering a specific antigen, many B cells change the isotype that they produce They stop making IgM and IgD, and now produce IgG, or IgA, or IgE

102
The Functional B Cell Receptor
Membrane-bound Igs are associated with two other proteins: Ig and Ig Ig and Ig have longer cytoplasmic tails than Ig and interact with intracellular signaling molecules

103
Affinity Maturation Thus somatic hypermutation results in mutant Igs on the surface of B cells Some of these mutants will have a higher affinity for the Ag - they will therefore be more likely to bind the Ag, and B cells bearing these molecules will be preferentially selected to mature into Ab-secreting cells As immune response proceeds, antibodies of higher affinity will be produced Called “affinity maturation”

104
Production of Membrane or Secreted Antibodies by Alternative RNA Processing

105
Switch Sequences or Switch Regions
Isotype switching - a recombination within the cluster of C genes that excises the previously expressed C gene and brings a different one into juxtaposition with the assembled V-region sequence Antigen specificity of the Ab remains unchanged, the isotype changes Flanking the 5’ side of each C gene, with the exception of the gene are highly repetitive sequences that mediate recombination

106
Each Human Immunoglobulin Isotype has Specialized Functions and Distinct Properties

Similar presentations