Download presentation
Presentation is loading. Please wait.

1
Izzati Adilah bt. Azmir 02-039
GENES TRANSFER Izzati Adilah bt. Azmir 02-039

2
Overview on Bacterial Gene Transfer
Bacteria are usually haploid Makes it easy to identify loss-of-function mutations in bacteria than in eukaryotes These usual recessive mutations are not masked by dominant genes in haploid species Bacteria reproduce asexually Therefore crosses are not used in the genetic analysis of bacterial species Rather, researchers rely on a similar phenomenon called genetic transfer In this process, a segment of bacterial DNA is transferred from one bacterium to another

3
Genetic transfer A process to transfer genetic material from a bacterium to another bacterium Enhances genetic diversity Confer resistance to antibiotic when one a antibiotic resistant bacterium transfer the gene to another bacterial cell

4
Mechanism of Gene Transfer
Conjugation Direct physical interaction between Donor and recipient cell Transduction When virus infects a bacterium and transfer genetic material Transformation Information is taken from a dead bacterium which releases it to the environment

5
Mechanisms of Gene Transfer

6
Conjugation

7
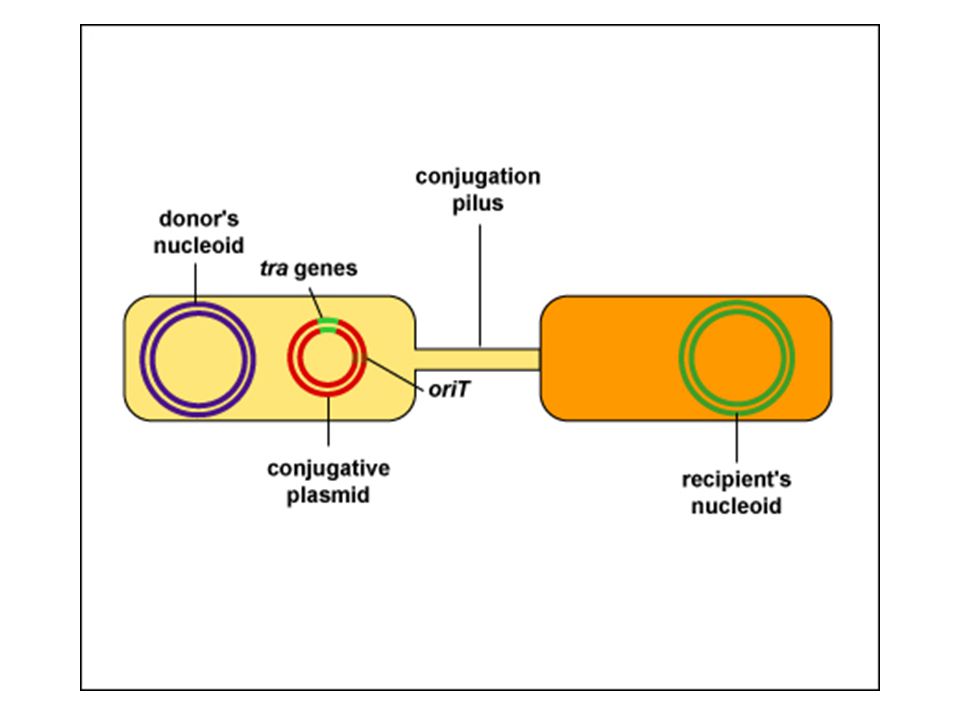
CONJUGATION Direct physical interaction between Donor and recipient cell E.g plasmid is transferred to a recipient cell from a donor Requires the presence of a special plasmid called the F plasmid.

8
Conjugation cont’d A “mating” process between a donor F+ (bacteria with fertility factor =plasmid) and an F- recipient cell. Occurs in Gram -ve enteric bacteria like E.coli Plasmids carry genes that are nonessential for the life of bacteria. Uses pili (sex pilus). E.g. plasmid replication enzymes. Causes medical Problem: R-Factor = antibiotic resistance! Enteric = gut bacteria
. E.g. plasmid replication enzymes. Causes medical Problem: R-Factor = antibiotic resistance! Enteric = gut bacteria.")
10
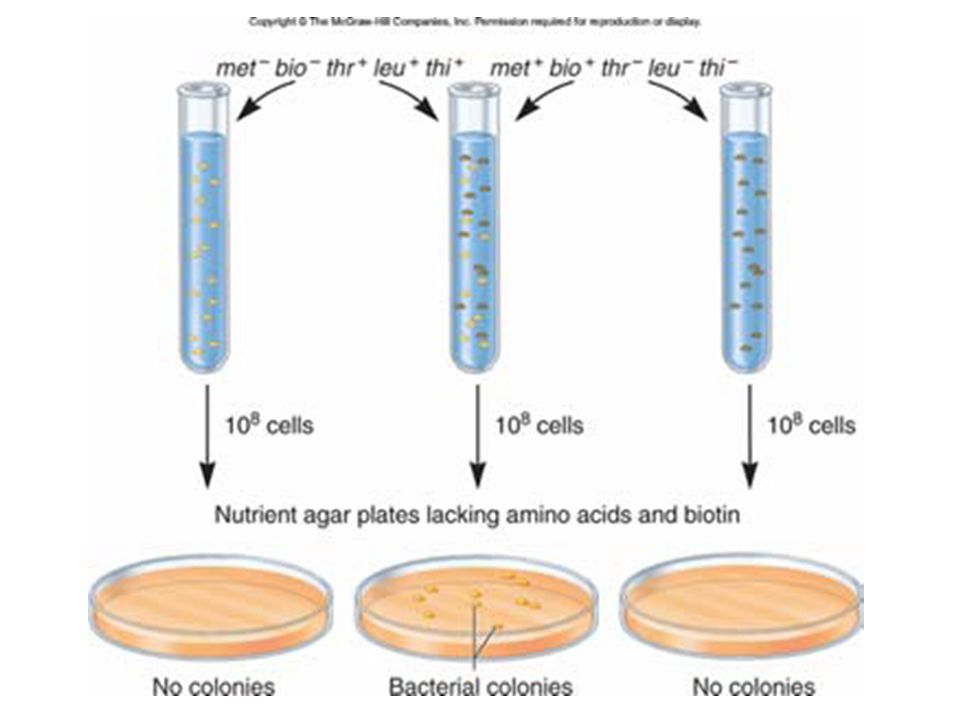
Discovery… Discovered in 1946 in bacteria by Joshua Lederberg and Edward Tatum They were studying strains of E. coli that had different nutritional growth requirements Auxotrophs cannot synthesize a needed nutrient Prototrophs make all their nutrients from basic components One auxotroph strain was designated bio– met– phe+ thr+ It required one vitamin (biotin) and one amino acid (methionine) It could produce the amino acids phenylalanine and threonine The other strain was designated bio+ met+ phe– thr–
 and one amino acid (methionine) It could produce the amino acids phenylalanine and threonine. The other strain was designated bio+ met+ phe– thr–")
12
The genotype of the bacterial cells that grew on the plates has to be bio+ met+ phe+ thr+
Lederberg and Tatum reasoned that some genetic material was transferred between the two strains Either the bio– met– phe+ thr+ strain got the ability to synthesize biotin and methionine (bio+ met+) Or the bio+ met+ phe– thr– strain got the ability to synthesize phenylalanine and threonine (phe+ thr+) The results of this experiment cannot distinguish between the two possibilities
 Or the bio+ met+ phe– thr– strain got the ability to synthesize phenylalanine and threonine (phe+ thr+) The results of this experiment cannot distinguish between the two possibilities.")
13
The need for physical contact
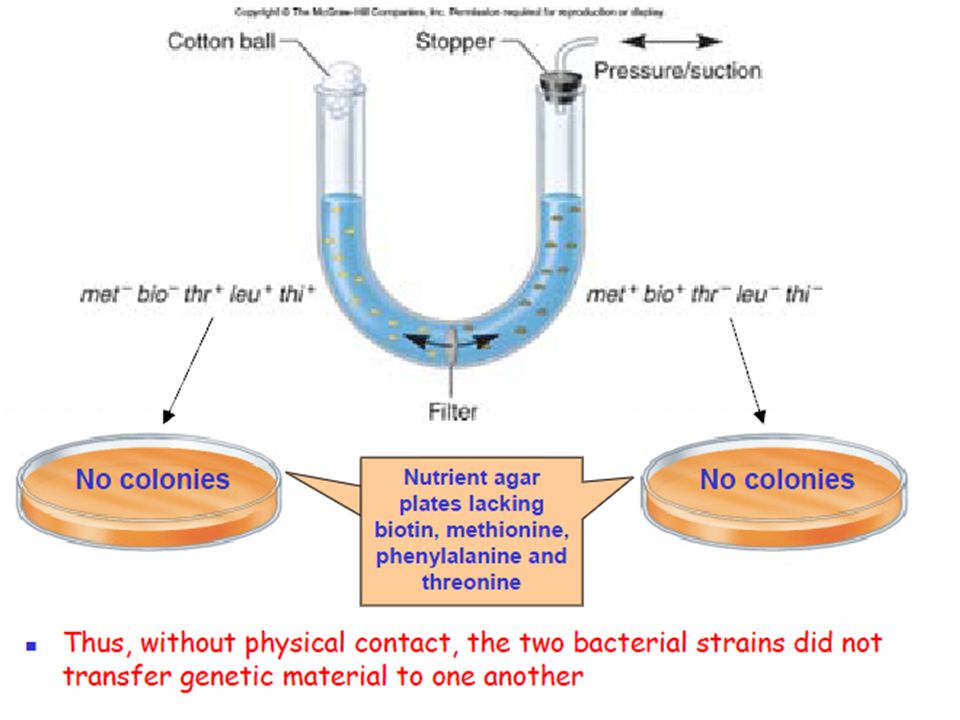
Bernard Davis later showed that the bacterial strains must make physical contact for transfer to occur He used an apparatus known as U-tube It contains at the bottom a filter which has pores that were large enough to allow the passage of the genetic material But small enough to prevent the passage of bacterial cells Davis placed the two strains in question on opposite sides of the filter Application of pressure or suction promoted the movement of liquid through the filter

15
Many species of bacteria can conjugate
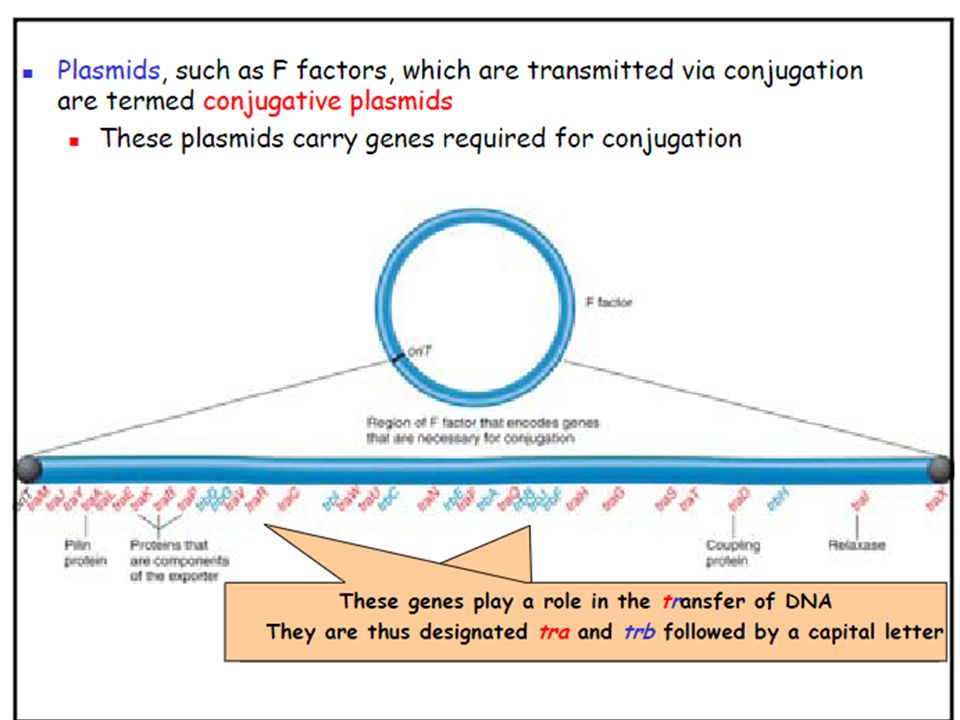
The term conjugation now refers to the transfer of DNA from one bacterium to another following direct cell-to cell contact Many species of bacteria can conjugate Only certain strains of a bacterium can act as donor cells Those strains contains a small circular piece of DNA termed the F factor (for Fertility factor) Strains containing the F factor are designated F+ Those lacking it are F– Plasmid is the general term used to describe extra-chromosomal DNA Plasmids, such as F factors, which are transmitted via conjugation are termed conjugative plasmids These plasmids carry genes required for conjugation
 Strains containing the F factor are designated F+ Those lacking it are F– Plasmid is the general term used to describe extra-chromosomal DNA. Plasmids, such as F factors, which are transmitted via conjugation are termed conjugative plasmids. These plasmids carry genes required for conjugation.")
17
Plasmid Small, circular pieces of DNA that are separated and replicated independently from the bacterial chromosome. Contains only a few genes that are usually not needed for growth and reproduction of the cell. But important in stressful situations F plasmid, facilitates conjugation Can give a bacterium new genes that may help for survival in changing environment. Some plasmids can integrate reversibly into the bacterial chromosome. An integrated plasmid is called an episome.

18
Plasmid There are several types of plasmids:
a. Conjugative plasmids – genes for sex pili and conjugation b. Dissimulation plasmids – genes for enzymes that catabolize unusual organic molecules (Pseudomonas species – toluene, camphor, petroleum products) c. Plasmids carrying genes for toxins or bacteriocins d. Plasmids carrying genes for resistance (R) factors i. Consist of two sets of genes – RTF (resistance transfer factor) and specific resistance genes (r-determinant)
 c. Plasmids carrying genes for toxins or bacteriocins. d. Plasmids carrying genes for resistance (R) factors. i. Consist of two sets of genes – RTF (resistance transfer factor) and specific resistance genes (r-determinant)")
19
Episome VS Plasmid Plasmid DNA is an extra-chromosomal DNA molecule, it cannot link up with chromosomal DNA, and it contains the genetic informations that are necessary for its own replication. Episomes is any kind of extra-chromosomal DNA that can link up with chromosomal DNA. That is the main difference between them two. Episomes are usually larger than other extra-chromosomal DNA. An example of episome are the viruses, because they intergrate their genetic material into the host's chromosomal DNA.

20
Mechanism of Conjugation
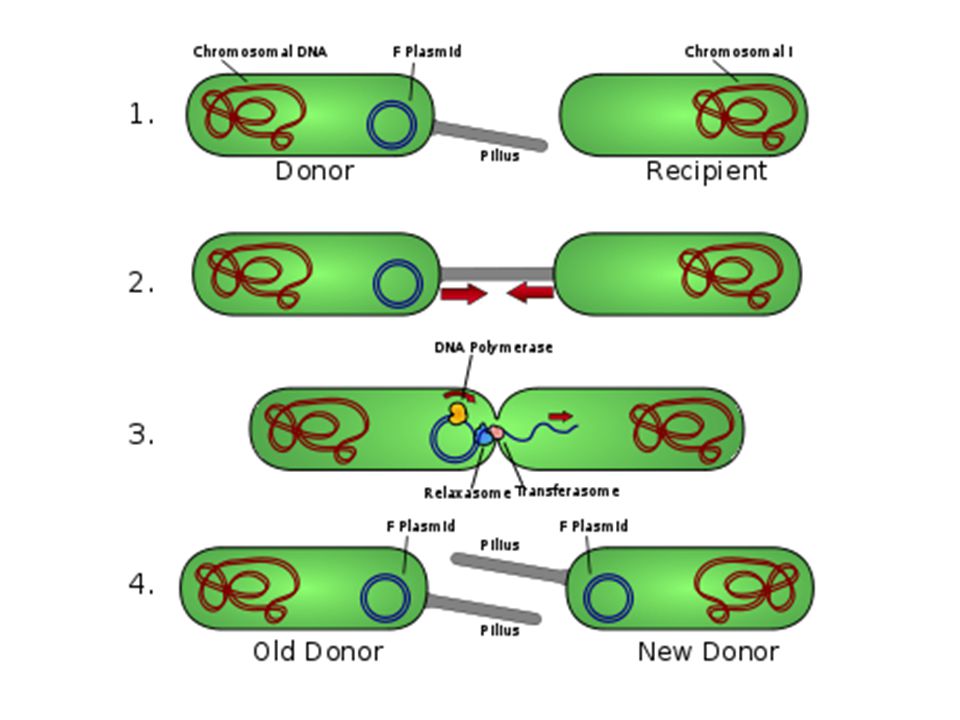
The first step in conjugation is the contact between donor and recipient cells This is mediated by sex pili (or F pili) which are made only by F+ strains These pili act as attachment sites for the F– bacteria Once contact is made, the pili shorten Donor and recipient cell are drawn closer together A conjugation bridge is formed between the two cells The successful contact stimulates the donor cells to begin the transfer process
 which are made only by F+ strains. These pili act as attachment sites for the F– bacteria. Once contact is made, the pili shorten. Donor and recipient cell are drawn closer together. A conjugation bridge is formed between the two cells. The successful contact stimulates the donor cells to begin the transfer process.")
22
F’ factors can be transferred through conjugation
The result of conjugation is that the recipient cell has acquired an F factor Thus, it is converted from an F– to an F+ cell The F+ cell remains unchanged In some cases, the F factor may carry genes that were once found on the bacterial chromosome These types of F factors are called F’ factors F’ factors can be transferred through conjugation This may introduce new genes into the recipient and thereby alter its genotype

23
Hfr Strains In the 1950s, Luca Cavalli-Sforza discovered a strain of E. coli that was very efficient at transferring chromosomal genes He designated this strain as Hfr (for High frequency of recombination) Hfr strains are derived from F+ strains
 Hfr strains are derived from F+ strains.")
24
Mechanism in Hfr Strains

26
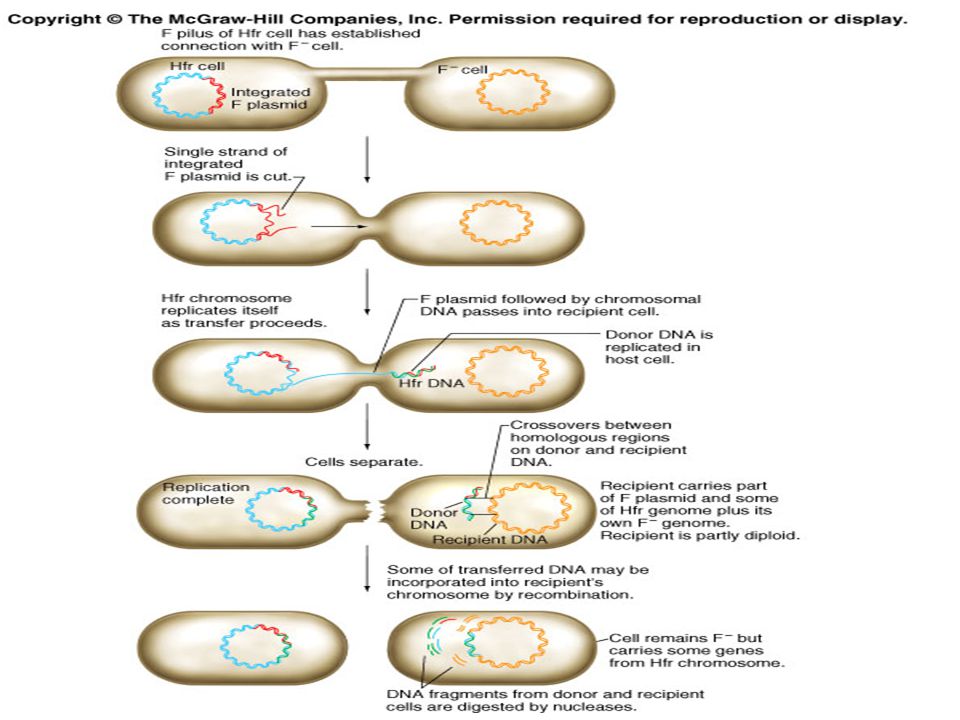
William Hayes demonstrated that conjugation between an Hfr and an F– strain involves the transfer of a portion of the Hfr bacterial chromosome The origin of transfer of the integrated F factor determines the starting point and direction of the transfer process The cut, or nicked site is the starting point that will enter the F– cell Then, a strand of bacterial DNA begins to enter in a linear manner

27
The F– cell does pick up chromosomal DNA
It generally takes about hours for the entire Hfr chromosome to be passed into the F– cell Most matings do not last that long Only a portion of the Hfr chromosome gets into the F– cell Since the nick is internal to the integrated F factor, only part of the plasmid is transferred and the F– cells does not become F+ The F– cell does pick up chromosomal DNA This DNA can recombine with the homologous region on the chromosome of the recipient cell This may provide the recipient cell with new combination of alleles

28
Hfr (High Frequency Recombination)
Hfr- bacterial plasmid integrates into the chromosome. Medical Problem: Hfr antibiotic resistance genes are passed during binary fission (every time the cell divides). Therefore, antibiotic resistance spreads very rapidly! When Hfr mate with F – bacteria, only the bacterial genes cross NOT plasmid genes. Genetic diversity results in this case due to recombination.
. Therefore, antibiotic resistance spreads very rapidly! When Hfr mate with F – bacteria, only the bacterial genes cross NOT plasmid genes. Genetic diversity results in this case due to recombination.")
30
Interrupted Mating Technique
Developed by Elie Wollman and François Jacob in the 1950s The rationale behind this mapping strategy The time it takes genes to enter the recipient cell is directly related to their order along the bacterial chromosome The Hfr chromosome is transferred linearly to the F– recipient cell Therefore, interrupted mating at different times would lead to various lengths being transferred The order of genes along the chromosome can be deduced by determining the genes transferred during short matings vs. those transferred during long matings

31
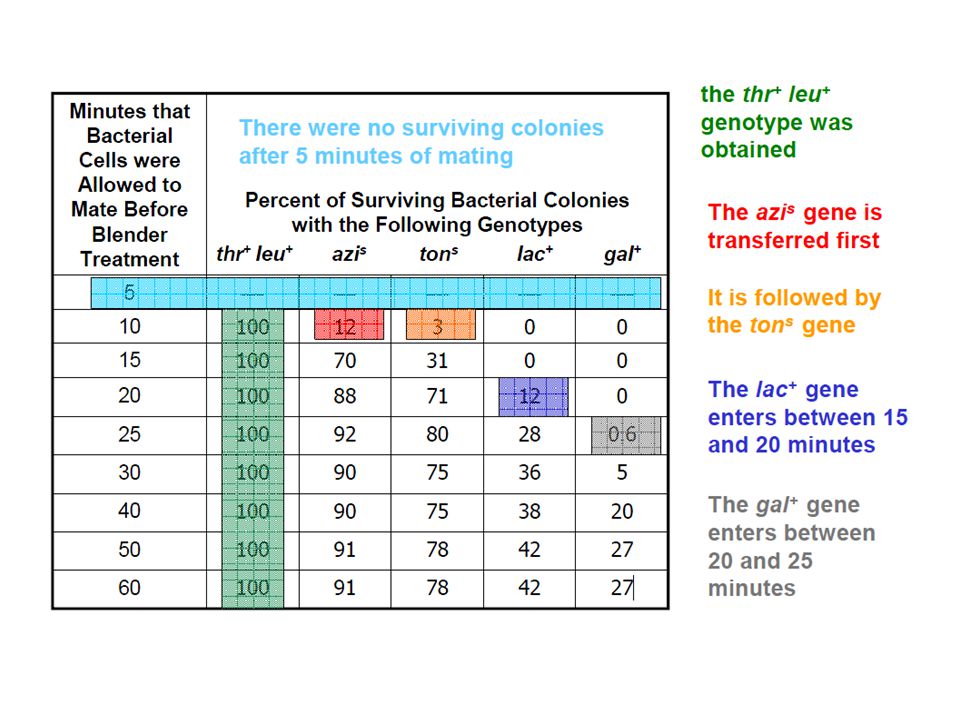
Wollman and Jacob started the experiment with two E. coli strains
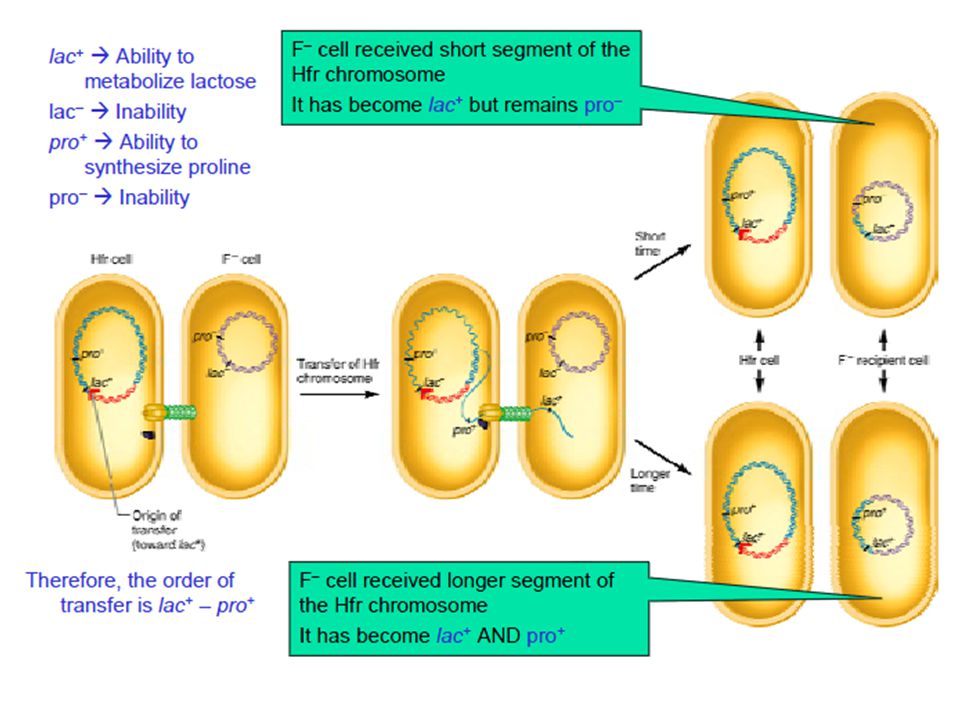
The donor (Hfr) strain had the following genetic composition thr+ : Able to synthesize the essential amino acid threonine leu + : Able to synthesize the essential amino acid leucine azis : Sensitive to killing by azide (a toxic chemical) tons : Sensitive to infection by T1 (a bacterial virus) strs : Sensitive to killing by streptomycin (an antibiotic) strr : Resistant to killing by streptomycin (an antibiotic) lac+ : Able to metabolize lactose and use it for growth gal + : Able to metabolize galactose and use it for growth The recipient (F–) strain had the opposite genotype thr– leu– azir tonr lac – gal – strr r = resistant
 strain had the following genetic composition. thr+ : Able to synthesize the essential amino acid threonine. leu + : Able to synthesize the essential amino acid leucine. azis : Sensitive to killing by azide (a toxic chemical) tons : Sensitive to infection by T1 (a bacterial virus) strs : Sensitive to killing by streptomycin (an antibiotic) strr : Resistant to killing by streptomycin (an antibiotic) lac+ : Able to metabolize lactose and use it for growth. gal + : Able to metabolize galactose and use it for growth. The recipient (F–) strain had the opposite genotype. thr– leu– azir tonr lac – gal – strr. r = resistant.")
32
Wollman and Jacob already knew that
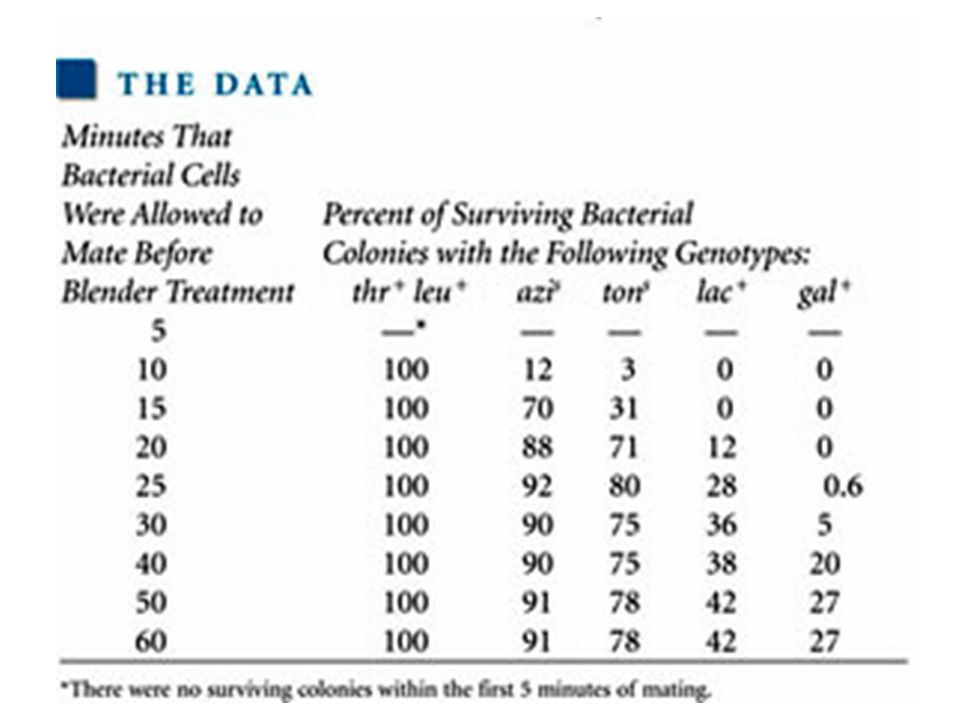
The thr+ and leu+ genes were transferred first, in that order Both were transferred within 5-10 minutes of mating Therefore their main goal was to determine the times at which genes azis, tons, lac+, and gal+ were transferred The transfer of the strs was not examined Streptomycin was used to kill the donor (Hfr) cell following conjugation The recipient (F– cell) is streptomycin resistant
 cell following conjugation. The recipient (F– cell) is streptomycin resistant.")
35
From these data, Wollman and Jacob constructed the following genetic map:
They also identified various Hfr strains in which the origin of transfer had been integrated at different places in the chromosome Comparison of the order of genes among these strains, demonstrated that the E. coli chromosome is circular

36
Summary

37
Conjugation Mechanism and its Product

38
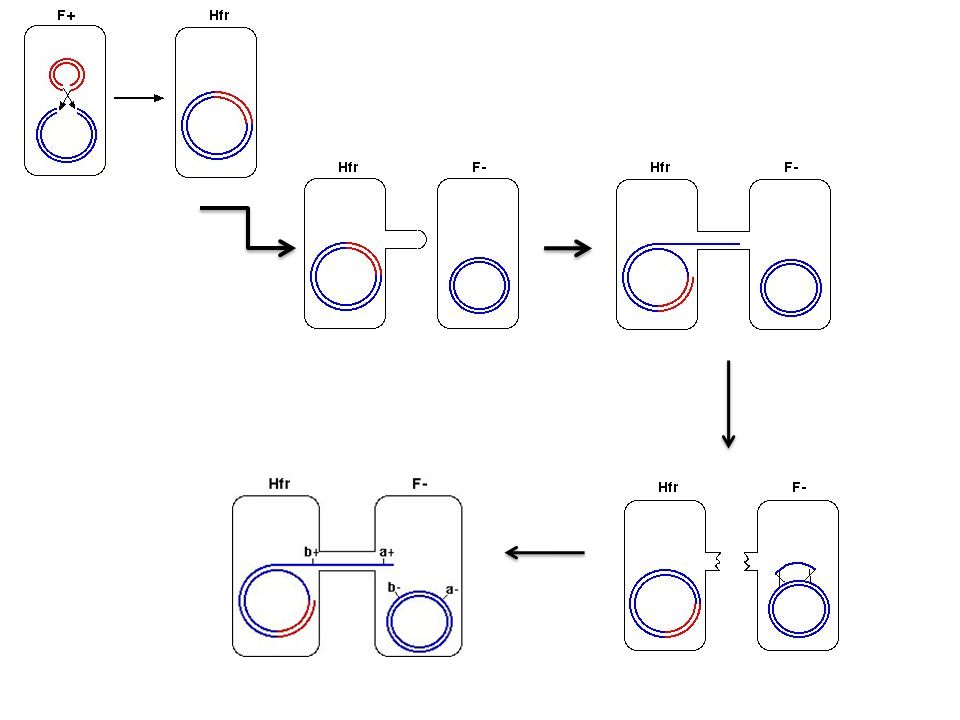
Basic conjugation (F+ and F-)
The difference F+ has a Fertility factor (F factor) The F factor is an episome that contains 19 genes and confers the ability to conjugate upon its host cell. From F+ cell to an F- cell, and the genetic material transferred is the F factor itself. The F+ cell initiates conjugation by extending an F pilus toward the F- cell. Among the genes present on the F factor are the genes encoding the proteins required for pilus construction.
 The F factor is an episome that contains 19 genes and confers the ability to conjugate upon its host cell. From F+ cell to an F- cell, and the genetic material transferred is the F factor itself. The F+ cell initiates conjugation by extending an F pilus toward the F- cell. Among the genes present on the F factor are the genes encoding the proteins required for pilus construction.")
39
Basic conjugation 1 2 3 4

40
Basic conjugation When finished, F pilus temporarily connects the two cells. F factor strand is nicked, and unwinding begins from the other strand. The nicked strand begins to transfer through the F pilus to the F- cell. As it does so, this strand begins to be replicated, as does circular strand remaining behind in the F+ cell.

41
Basic conjugation 1 2 3 4

42
Basic conjugation cont’d
Eventually, the nicked strand completely passes through to the recipient cell, and is completely replicated. Produces a new F factor in the recipient cell. Pilus is broken, severing the connection between the two cells. Both cells now contain an F factor, become F+. New F+ cell (which was the F- cell), can now initiate conjugation with another F- cell.
, can now initiate conjugation with another F- cell.")
43
Conjugation Involving Hfr Bacteria
Occasionally, the F factor integrates into a random position in the bacterial chromosome. When this happens, the bacterial cell is called Hfr instead of F+. Hfr bacteria are still able to initiate conjugation with F- cells, but the outcome is completely different from conjugation involving F+ bacteria:

44
Hfr cells When F factor integrates into the bacterial chromosome at a random location. When DNA transfer begins, the Hfr cell tries to transfer the entire bacterial chromosome to the F- cell. The first DNA to be transferred is chromosomal DNA, and the last DNA to be transferred will be the F factor DNA. Transfer of the bacterial chromosome is almost never complete. Pili tend to break, disrupting DNA transfer before the entire chromosome can be transferred. So, F factor almost never transferred to the recipient cell & remains F-. But this cell will receive new DNA from the Hfr and new DNA can undergo recombination at a high frequency with the host chromosome.

46
F' Conjugation to F- F factors can occasionally integrate into the bacterial chromosome (producing an Hfr cell from an F+ cell). Integrated F factors can occasionally excise themselves from the bacterial chromosome. If this excision occurs properly, the Hfr cell becomes an F+ again. But the excision is sometimes poor. F factor takes a small segment of the bacterial chromosome with it. So some of the chromosomal DNA become associated with the episome. When this happens, the cell is called an F'. Plasmid DNA is an extra-chromosomal DNA molecule, it cannot link up with chromosomal DNA, and it contains the genetic informations that are necessary for its own replication. Episomes is any kind of extra-chromosomal DNA that can link up with chromosomal DNA. That is the main difference between them two. Episomes are usually larger than other extra-chromosomal DNA. An example of episome are the viruses, because they intergrate their genetic material into the host's chromosomal DNA

47
Conjugation involving F' cells allows for the possibility of recombination, as shown below:
When F factor begins to transfer its DNA to the recipient cell, it will transfer the small segment of chromosomal DNA as well. F' cells are able to initiate conjugation with F- cells because of the presence of the F factor. Just as in the F+/F- mating, both cells wind up with a copy of the episome. The cell that was F- now has the F factor and is now F'.

48
F' Conjugation to F- The F' cell has a full complement of chromosomal genes; however, some of those genes are now on the episome. F' cells are able to initiate conjugation with F- cells because of the presence of the F factor. In the transfer, it will transfer the small segment of chromosomal DNA as well. Just as in the F+/F- mating, both cells wind up with a copy of the episome. The cell that was F- now has the F factor (along with the piece of chromosomal DNA) and is therefore now F'. This cell, however, also has a complete chromosome, so it will be diploid for the segment of chromosomal DNA on the episome. Such a partially diploid bacterial cell is called a merozygote. The chromosomal DNA on the episome can undergo recombination at high frequency with its homologous sequence on the chromosome.
 and is therefore now F . This cell, however, also has a complete chromosome, so it will be diploid for the segment of chromosomal DNA on the episome. Such a partially diploid bacterial cell is called a merozygote. The chromosomal DNA on the episome can undergo recombination at high frequency with its homologous sequence on the chromosome.")
49
Let's summarize F+ mating with F- produces 2 F+
Hfr mating with F- produces Hfr and F- F' mating with F- produces F' and F' merozygote (an incomplete bacterial zygote having only a fragment of the genome from one of the two parent cells)
")
50
Quiz What happens when: F+ meets F- Hfr meets F- F’ meets F-
What is F’ by the way? What is Hfr and how did it occur?

51
Transduction

52
TRANSDUCTION The transfer of genetic material from donor bacteria to recipient bacteria via transducing agent (bacterial viruses called bacteriophage). Discovered in 1952 by Zinder & Lederberg. Two kinds of transduction: generalized and specialized.
. Discovered in 1952 by Zinder & Lederberg. Two kinds of transduction: generalized and. specialized.")
53
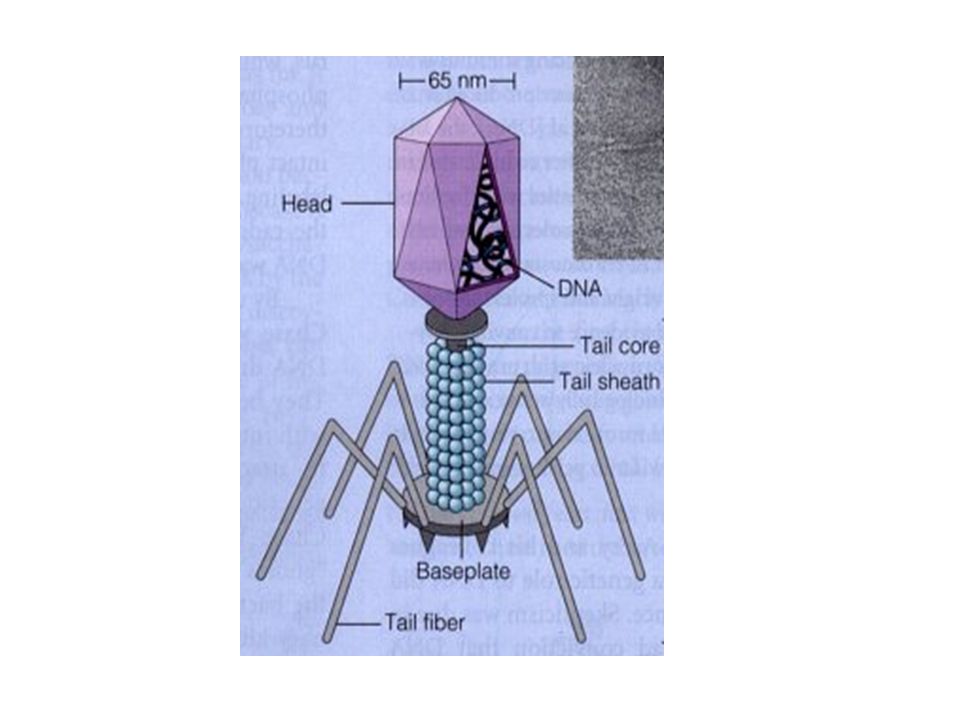
Transduction A bacteriophage is a virus that specifically attacks bacterial cells It is composed of genetic material surrounded by a protein coat It can undergo two types of cycles Lytic Lysogenic

55
It will switch to the lytic cycle Prophage can exist in a dormant state for a long time Virulent phages only undergo a lytic cycle Temperate phages can follow both cycles

56
Transduction Phages that can transfer bacterial DNA include
P22, which infects Salmonella typhimurium P1, which infects Escherichia coli Both are temperate phages

57
Generalized transduction
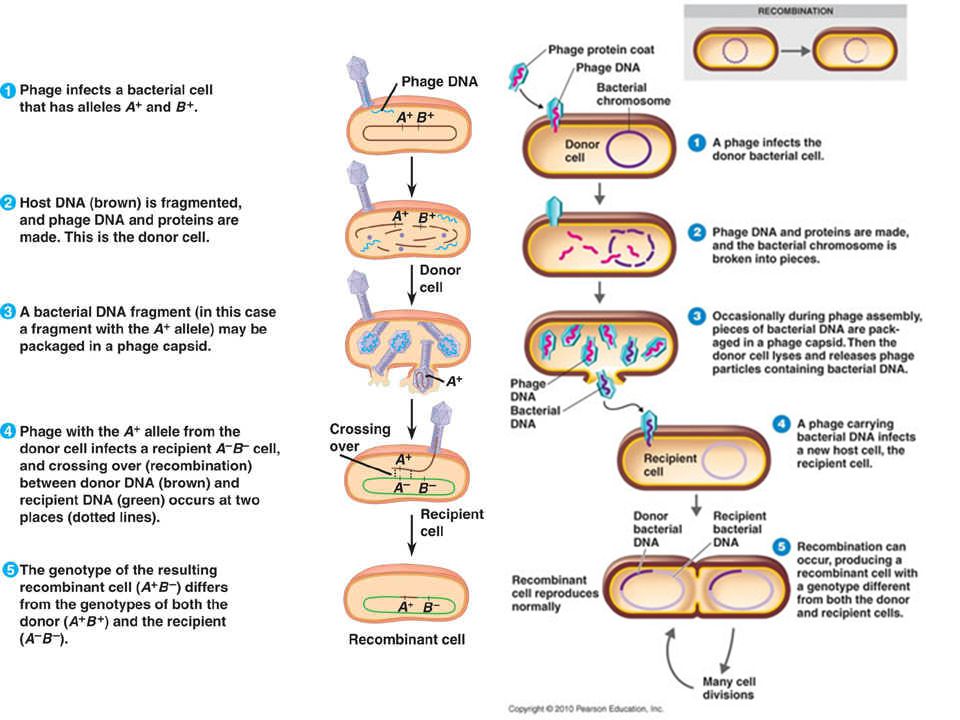
Starts with the LYTIC CYCLE where a T- even phage infects E. coli killing the host cell, and synthesizing 2,000 copies of itself. The T-even phage randomly packages bacterial DNA in a few defective phages. Once a T – even phage infects another E. coli, this genetic information can be recombined into the host cell without causing the lytic cycle. New genetic information is thereby transduced from one bacteria to another.

58
Generalized Transduction

59
Generalized Transduction

60
Specialized Transduction
Lambda phage infects E.coli but does not lyse the cell immediately. Instead it integrates into chromosome of the bacteria as a prophage and remains dormant. This is called the LYSOGENIC CYCLE. Phage genes are replicated and passed to all daughter cells until the bacteria is under environmental stress, from lack of nutrients, etc. Then phage gene will excise from the nucleoid and enter the LYTIC CYLE taking one adjacent gene for galactose metabolism.

61
It will switch to the lytic cycle Prophage can exist in a dormant state for a long time Virulent phages only undergo a lytic cycle Temperate phages can follow both cycles

62
Specialized Transduction cont’d
The gal transducing phage (lambda) makes ~ 2,000 copies of itself with the gal gene, and infects other E.coli. When gal integrates into the nucleoid of other E. coli, it may provide these bacteria with a new capacity to metabolize galactose.
 makes ~ 2,000 copies of itself with the gal gene, and infects other E.coli. When gal integrates into the nucleoid of other E. coli, it may provide these bacteria with a new capacity to metabolize galactose.")
63
Specialized Transduction Graphic

64
Comparison of Bacteriophage
Comparison of bacteriophage transduction in E.coli. Generalized Specialized T even phage lambda phage lytic cycle lysogenic random packaging specific gal gene

66
Transformation

67
TRANSFORMATION The passage of homologous DNA from a dead donor cell to a living recipient cell. Occurs in Streptococcus pneumoniae. When S. pneumo dies the DNA can be absorbed by a living S. pneumo and recombined into the chromosome. The gene for capsule formation is obtained in this way, as is a gene for penicillin resistance. Discovered in 1929 by Fredrick Griffith.

68
Griffith’s Transformation Experiment

69
Griffith’s experiment

70
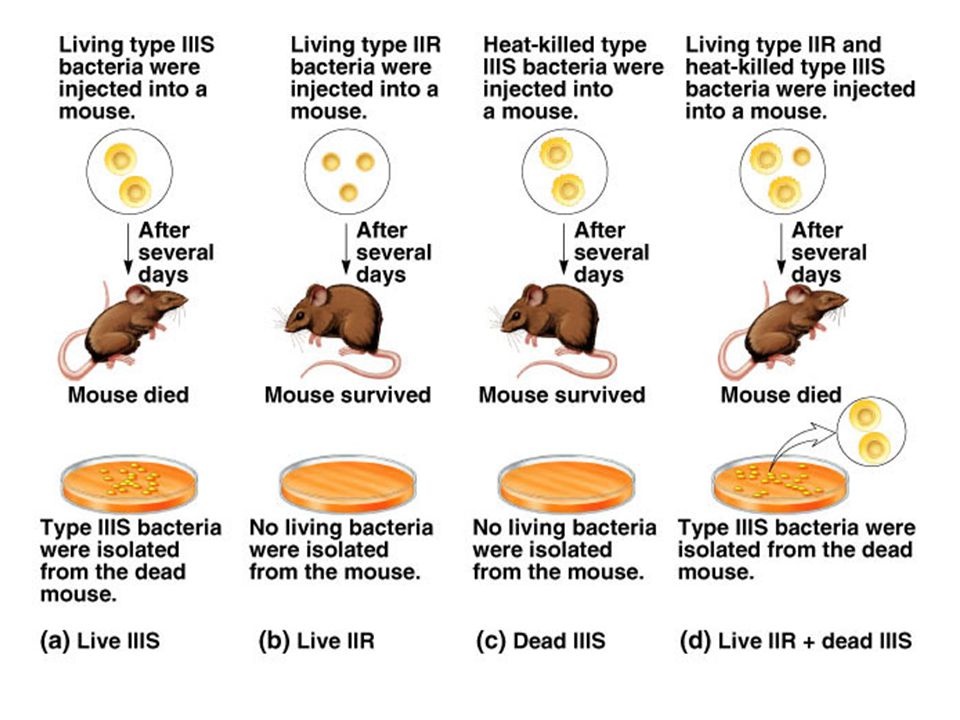
(a) Inject living encapsulated bacteria into mice, mice die, encapsulated bacteria isolated from dead mice. (b) Inject living nonencapsulated bacteria into mice, mice remain healthy, a few non-encapsulated bacteria can be isolated from the living mice – most phagocytized by leukocytes. (c) Inject heat-killed encapsulated bacteria into mice, mice remain healthy, no bacteria isolated from the living mice. (d) Inject living non-encapsulated and heat-killed encapsulated bacteria into mice, mice die, isolated encapsulated bacteria from dead mice.
 Inject living nonencapsulated bacteria into mice, mice remain healthy, a few non-encapsulated bacteria can be isolated from the living mice – most phagocytized by leukocytes. (c) Inject heat-killed encapsulated bacteria into mice, mice remain healthy, no bacteria isolated from the living mice. (d) Inject living non-encapsulated and heat-killed encapsulated bacteria into mice, mice die, isolated encapsulated bacteria from dead mice.")
72
The Experiments of Avery, MacLeod and McCarty
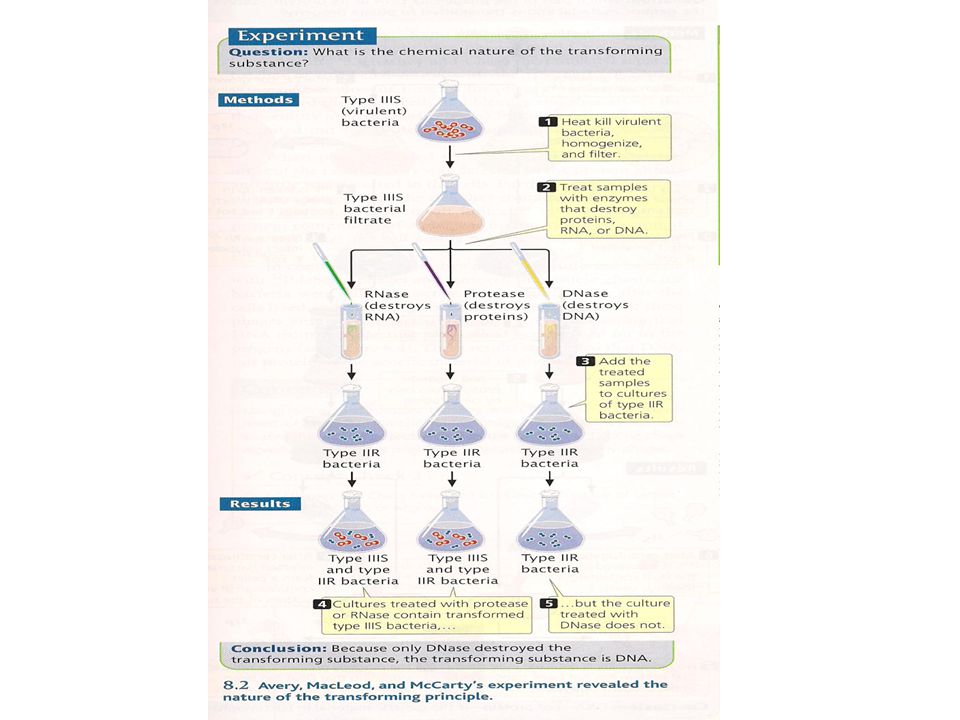
Avery, MacLeod and McCarty realized that Griffith’s observations could be used to identify the genetic material They carried out their experiments in the 1940s At that time, it was known that DNA, RNA, proteins and carbohydrates are major constituents of living cells They prepared cell extracts from type IIIS cells containing each of these macromolecules Only the extract that contained purified DNA was able to convert type IIR into type IIIS

74
Hershey and Chase Experiment with Bacteriophage T2
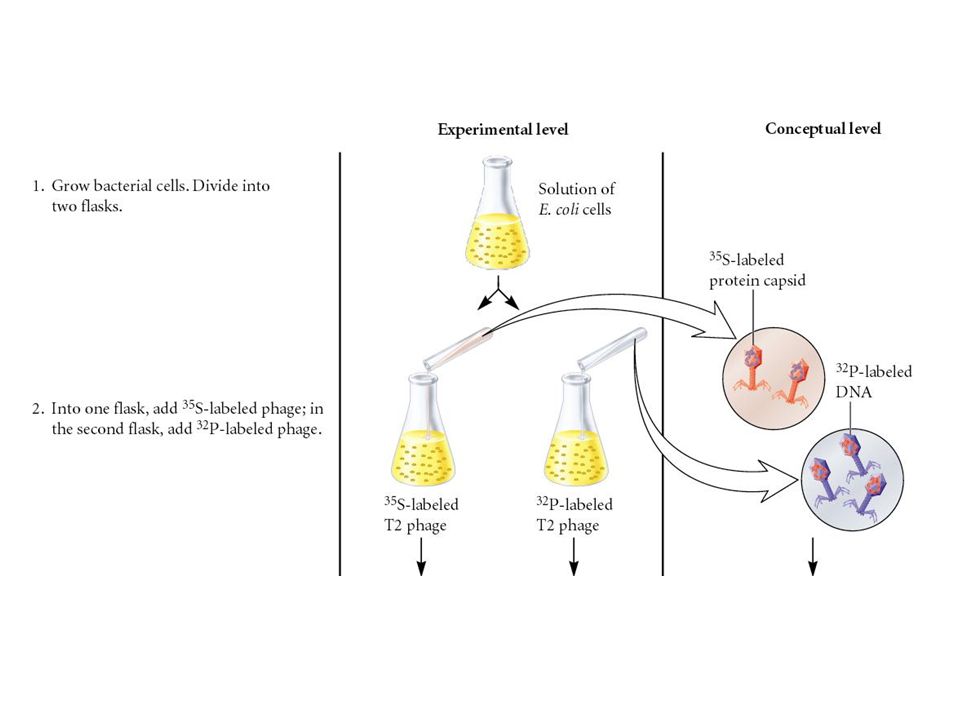
In 1952, Alfred Hershey and Marsha Chase provided further evidence that DNA is the genetic material Inside the capsid They studied the bacteriophage T2 It is relatively simple since its composed of only two macromolecules DNA and protein Made up of protein

75
Life cycle of the T2 bacteriophage

76
The Hershey and Chase experiment can be summarized as follows:
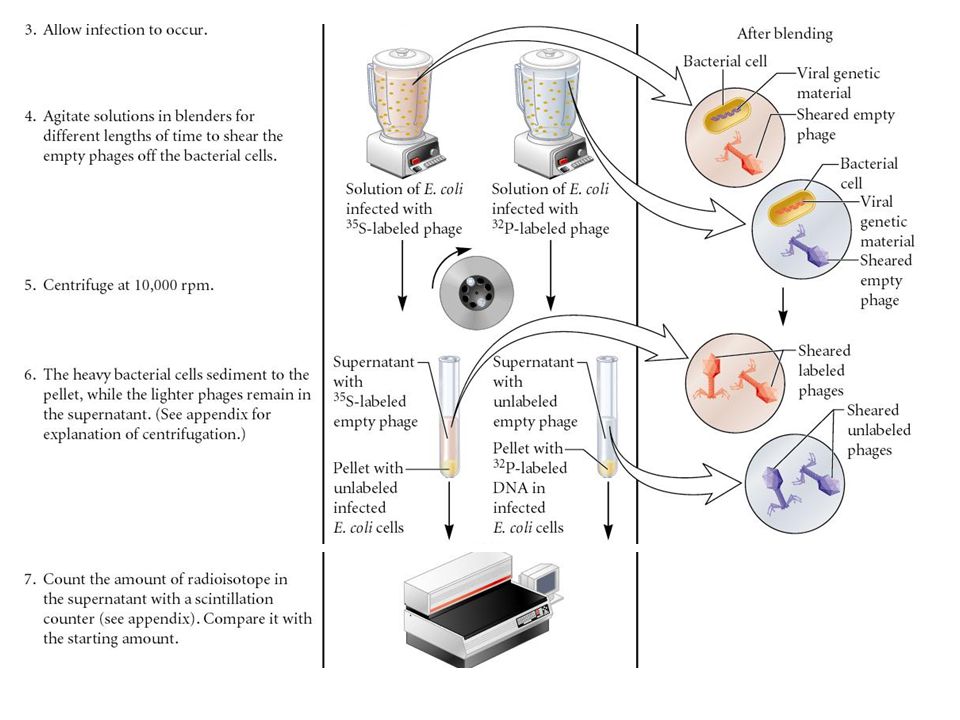
Used radioisotopes to distinguish DNA from proteins 32P labels DNA specifically 35S labels protein specifically Radioactively-labeled phages were used to infect nonradioactive Escherichia coli cells After allowing sufficient time for infection to proceed, the residual phage particles were sheared off the cells => Phage ghosts and E. coli cells were separated Radioactivity was monitored using a scintillation counter

79
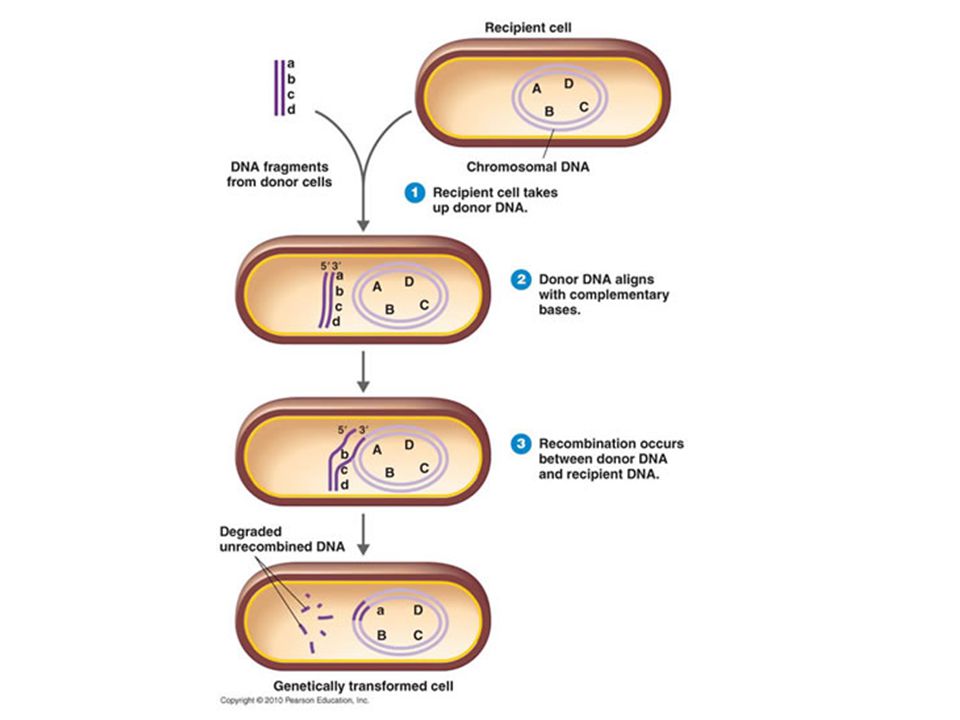
Transformation The process by which a bacterium will take up extracellular DNA released by a dead bacterium It was discovered by Frederick Griffith in 1928 while working with strains of Streptococcus pneumoniae There are two types Natural transformation DNA uptake occurs without outside help Artificial transformation DNA uptake occurs with the help of special techniques

81
Transformation Graphic

82
TRANSPOSITION Transposons (jumping genes) are big chunks of DNA that randomly excise and relocate on the chromosome. Transposons were discovered in 1950 by Barbara McLintock in corn. Causes antibiotic resistance in Staph. aureus, the famous methicillin resistant Staphlococcus aureus (MRSA) strain!
 strain!")
Similar presentations












 Lecture 10 Microbial Genetics (Text Chapter: 10.1-10.4; 10.6-10.14)>")






 2. How do bacteria replicate?>")