Download presentation
Presentation is loading. Please wait.

1
Chapter 7: DNA Repair Pathways

2
We totally missed the possible role of enzymes in DNA repair…
We totally missed the possible role of enzymes in DNA repair…. I later came to realize that DNA is so precious that probably many distinct repair mechanisms would exist. Nowadays one could hardly discuss mutation without considering repair at the same time. Francis Crick, Nature (1974), 248:766
, 248:766.")
3
7.1 Introduction

4
DNA damage poses a continuous threat to genomic integrity.
Cells have evolved a range of DNA repair enzymes and repair polymerases as complex as the DNA replication apparatus itself. DNA replication, repair, and recombination share many common features.

5
7.2 Mutations and DNA damage

6
Spontaneous mutations
Occur as a result of natural processes in cells. e.g. DNA replication errors Induced mutations Occur as a result of interaction of DNA with an outside agent that causes DNA damage.

7
Mutations are of fundamental importance
Mutations are important as the major source of genetic variation that drives evolutionary change. Mutations may have deleterious or (rarely) advantageous consequences to an organism or its descendents. Mutant organisms are important tools for molecular biologists in characterizing the genes involved in cellular processes.
 advantageous consequences to an organism or its descendents. Mutant organisms are important tools for molecular biologists in characterizing the genes involved in cellular processes.")
8
The simplest type of mutation is a nucleotide substitution.
Mutations that alter a single nucleotide are called point mutations.

9
Transitions and transversions can lead to silent, missense, or nonsense mutations
Transition mutations replace one pyrimidine base with another, or one purine base with another. Transversion mutations replace a pyrimidine with a purine or vice versa. In humans, the ratio of transitions to transversions is approximately 2:1

10
A transition or transversion mutation can be permanently incorporated by DNA replication.

12
Whether or not nucleotide substitutions have a phenotypic effect depends on:
Do they alter a critical nucleotide in a gene regulatory region? Do they alter a critical nucleotide in the template for a functional RNA molecule? Are they silent, missense, or nonsense mutations in a protein-coding gene?

13
Silent mutations Mutations that change the nucleotide sequence without changing the amino acid sequence are called synonymous mutations or silent mutations.

14
Missense mutations Nucleotide substitutions in protein-coding regions that do result in changed amino acids are called nonsynonymous mutations or missense mutations. May alter the biological properties of the protein. Sickle cell anemia is an AT→TA transversion: Glutamic acid codon in the -globin gene replaced by a valine codon

15
Nonsense mutations A nucleotide substitution that creates a new stop codon is called a nonsense mutation. Causes premature chain termination during protein synthesis. Nearly always a nonfunctional product.

16
Insertions or deletions can cause frameshift mutations
If the length of an insertion or deletion is not an exact multiple of three nucleotides, this results in a shift in the reading frame of the resulting mRNA. Usually leads to production of a nonfunctional protein.

18
Expansion of trinucleotide repeats leads to genetic instability
Trinucleotide repeats can adopt triple helix conformations and unusual DNA secondary structures that interfere with transcription and DNA replication. Expansion of trinucleotide repeats leads to certain genetic neurological disorders.

19
Repeat expansion can occur by two different mechanisms:
Unequal crossing over. Slippage during DNA replication.

20
Unequal crossing over A trinucleotide repeat in one chromosome misaligns for recombination during meiosis with a different copy of the repeat in the homologous chromosome. Recombination increases the number of repeats on one chromosome, resulting in a duplication. On the other chromosome, there is a deletion.

21
Slippage during DNA replication
During DNA replication the DNA melts and then reanneals incorrectly in the repeated region, resulting in re-replication of an additional repeat.

22
General classes of DNA damage
Spontaneous damage to DNA can occur through the action of water in the aqueous environment of the cell. A mutagen is any chemical agent that causes an increase in the rate of mutation above the spontaneous background.

23
Three general classes of DNA damage
Single base changes Structural distortion DNA backbone damage

24
Single base changes A single base change or “conversion” affects the DNA sequence but has only a minor effect on overall structure. Deamination is the most frequent and important kind of hydrolytic damage. Methylated cytosines are “hotspots” for spontaneous mutation in vertebrate DNA because deamination of 5-methylcytosine generates thymine.

25
Alkylating agents such as nitrosamines lead to the formation of O6-methylguanosine.
This modified base often mispairs with thymine. Can result in a GC→GT→AT point mutation after DNA replication.

26
Oxidizing agents generated by ionizing radiation and chemicals that generate free radicals can lead to formation of 8-oxoguanine (oxoG) OxoG can form a Hoogsteen base pair with adenine. Gives rise to a GC→TA transversion. One of the most common mutations found in human cancers.

28
Structural distortion
UV radiation induces that formation of a cyclobutane ring between adjacent thymines, forming a T-T dimer. The T-T dimer distorts the double helix and can block transcription and replication. UV radiation can also induce dimers between cytosine and thymine.

29
Other bulky adducts can be induced by chemical mutagenesis.
Structural distortion can be caused by intercalating agents and base analogs: Ethidium bromide has several flat polycyclic rings that insert between the DNA bases. 5-bromouracil, an analog of thymine, can mispair with guanine.

30
DNA backbone damage Formation of abasic sites
Loss of the nitrogenous base from a nucleotide. Generated spontaneously by the formation of unstable base adducts. Double-stranded DNA breaks Induced by ionizing radiation and a wide range of chemical compounds. The most severe type of DNA damage.

31
Cellular responses to DNA damage
Damage bypass Damage reversal Damage removal

33
Lesion bypass

34
Translesion synthesis (TLS)
Specialized low-fidelity, “error-prone” DNA polymerases transiently replace the replicative polymerases and copy past damaged DNA. Typical error rates range from 10-1 to 10-3 per base pair.

35
Error-prone DNA polymerases
May insert incorrect nucleotides opposite the lesion: nucleotide substitution May skip past and insert correct nucleotides opposite bases downstream: frameshift A trade-off between death and a risk of high mutation rate.

38
DNA polymerase eta () Performs translesion synthesis past TT dimers by inserting AA. Has an extra wide active site that can accommodate two dNTPs instead of one. Van der Waals forces and hydrogen-bonding interactions hold the TT dimer so that the two thymines can be paired with two adenines.

39
Direct reversal of DNA damage

40
Reversal of thymine-thymine dimers by DNA photolyase
In most organisms, UV radiation damage to DNA can be directly repaired. DNA photolyase uses energy from near UV to blue light to break the covalent bonds holding two adjacent pyrimidines together.

41
A B

42
DNA photolyase has two cofactors:
A pigment that absorbs UV/blue light Fully reduced flavin dinucleotide (FADH-) Splitting of the TT dimer is initiated by an electron transferred from photoexcited FADH- to the TT dimer already bound to the enzyme.
 Splitting of the TT dimer is initiated by an electron transferred from photoexcited FADH- to the TT dimer already bound to the enzyme.")
43
The TT dimer is flipped out of the DNA helix and brought very close to FADH-.
An electron is transferred from FADH- and the dimer is split. The electron is then returned to the transiently formed flavin radical in less than a nanosecond.

44
Photolyases are an ancient and efficient means of repairing UV-damaged DNA.
Placental mammals including humans, however, do not have a photoreactivation pathway.

45
Damage reversal by DNA methyltransferase
Methyltransferase catalyzes the transfer of the methyl group on O6-methylguanine to the sulfhydryl group of a cysteine residue on the enzyme.

46
Damage reversal by DNA methyltransferase
DNA methyltransferase binds the minor groove of the DNA. The minor groove widens and the DNA bends by 15° away from the enzyme. The O6-methylguanine flips out from the double helix into the active site. A sulfhydryl group of a cysteine in the active site accepts the methyl group from guanine.

47
Does DNA methyltransferase fit the classic definition of an enzyme?

48
Repair of single base changes and structural distortions by removal of DNA damage

49
Multiple dynamic protein interactions are involved in all repair processes.
Ordered hand-off of damaged DNA from one protein or protein complex to another. DNA repair proteins are modular.

50
The repair machinery must gain access to the DNA
Upon sensing DNA damage, nucleosomes are disassembled by histone modification and chromatin remodeling. After repair, PCNA recruits chromatin assembly factors to restore nucleosomes.

51
Pathways for repair of single base changes and structural distortion
Base excision repair Mismatch repair Structural distortion Nucleotide excision repair

52
Base excision repair The correction of single base changes that are due to conversion of one base to another. Specific DNA glycosylases recognize and excise the damaged base. How do DNA repair proteins find the rare sites of damage in a vast expanse of undamaged DNA?

53
A series of “gates” within the hOGG1 enzyme
Model for DNA damage recognition by 8-oxoguanine DNA glycosylase 1 (hOGG1) A series of “gates” within the hOGG1 enzyme hOGG1 first binds nonspecifically to DNA.
 A series of gates within the hOGG1 enzyme. hOGG1 first binds nonspecifically to DNA.")
54
If the enzyme encounters a normal GC base pair, then:
The G is transiently extruded into a G-specific pocket and returned to the double helix. If the enzyme encounters a oxoG-C base pair, then: The oxoG is extruded into the G-specific pocket and then inserted into a lesion recognition pocket where it is excised.

56
Base excision repair pathway in mammalian cells
A DNA glycosylase recognizes and excises the damaged base. An endonuclease cleaves the phosphodiester bond either 3′ or 5′ of the abasic site. An endonuclease removes 1-10 nucleotides. DNA polymerase replaces the missing nucleotides. DNA ligase seals the gap.

58
Mismatch repair The correction of mismatched base pairs which result from DNA polymerase errors during replication. A large region of DNA including the mismatch is excised. The method of strand discrimination in mammalian cells is currently unknown.

59
Hereditary nonpolyposis colorectal cancer: a defect in mismatch repair
3 to 5% of all colorectal cancers Inherit one inactive mismatch repair allele. Somatic loss of wild-type allele. Defective mismatch repair mechanism. Accumulation of mistakes during DNA replication. Microsatellite instability. 80% lifetime risk of developing colorectal cancer.

60
Mismatch repair pathway in mammalian cells:
Damage recognition by the MutS/MutL complex. One model proposes that MutS /MutL then diffuses for several thousand nucleotides either 5′ or 3′.

61
A 5′ or 3′ single-strand break is generated by EXO1 in association with PCNA and RFC.
5′→3′ or 3′→5′ progessive exonuclease activity of EXO1 removes the mismatch. 5′→3′ repair synthesis is mediated by DNA polymerase and associated factors. Ligation of the remaining gap is catalyzed by DNA ligase I.

63
Recurrent theme in DNA repair
Hand-off of damaged DNA from a complex with nuclease activity to a complex with polymerase activity to a complex with ligase activity.

65
Reconstitution of mismatch repair in an in vitro system showed that:
The recognition protein MutS, the exonuclease EXO1 and DNA polymerase are indispensible for repair.

66
Nucleotide excision repair
Repair of structural distortion e.g. bulges from thymine-thymine dimers induced by UV irradiation. Global genome repair (GGR) pathway: repair of lesions in the whole genome. Transcription coupled repair (TCR) pathway: repair of lesions in the transcribed strand of active genes.
 pathway: repair of lesions in the whole genome. Transcription coupled repair (TCR) pathway: repair of lesions in the transcribed strand of active genes.")
67
Mammalian nucleotide excision repair pathway
Damage recognition by the cooperative binding of XPC, RPA, XPA, and TFIIH. XPC binds first, followed by the other proteins. TFIIH is a multiprotein complex that also plays an important role in gene transcription.

68
Unwinding of the duplex DNA is promoted by the action of XPB and XPD helicases, which are subunits of the TFIIH complex. The endonuclease XPG makes a 3′ incision, and a 5′ incision is made by the endonuclease XPF-ERCC1. The damaged strand is released (24 to 32 nt).
.")
69
Repair synthesis is mediated by DNA polymerase or .
Ligation of the remaining gap in the DNA backbone is mediated by DNA ligase I.

71
Xeroderma pigmentosum and related disorders: defects in nucleotide excision repair
Autosomal recessive disorder. Photosensitivity. Greatly increased risk of sunlight-induced skin cancer. Neurological degeneration. Defects in nucleotide excision repair or in T-T dimer translesion synthesis.

72
TFIIH complex (with XPB, XPD helicases)
Which of the repair pathway components are defective in xeroderma pigmentosum? XPA RPA XPG XPC TFIIH complex (with XPB, XPD helicases) XPF/ERCC1 PCNA RFC DNA polymerase / DNA ligase I DNA polymerase
 XPF/ERCC1. PCNA. RFC. DNA polymerase / DNA ligase I. DNA polymerase ")
73
Xeroderma pigmentosum complementation groups
Seven complementation groups (XPA to XPG) and xeroderma pigmentosum variant (XPV). Complementation group: When fibroblast cells of two different patients with the same defect are fused in vitro, the DNA damage is not repaired.
 and xeroderma pigmentosum variant (XPV). Complementation group: When fibroblast cells of two different patients with the same defect are fused in vitro, the DNA damage is not repaired.")
74
Nucleotide excision repair is determined by the uptake of radiolabeled thymidine into DNA.
If the two patients have different gene defects, the cells correct each other and the DNA damage is repaired.

75
Treating xeroderma pigmentosum
Early diagnosis and light-protective lifestyle. Topical application of DNA repair enzymes.

76
Two other nucleotide excision repair deficiency syndromes
Trichothiodystrophy Cockayne syndrome

77
7.6 Double-strand break repair by removal of DNA damage

78
Double-strand breaks in DNA are induced by reactive oxygen species, ionizing radiation, and chemicals the generate reactive oxygen species (free radicals). Repaired by homologous recombination or nonhomologous end-joining.

79
Homologous recombination
Repairs double-strand breaks by retrieving genetic information from an undamaged homologous chromosome. Nonhomologous end-joining (NHEJ) Rejoins double-strand breaks via direct ligation of the DNA ends without any requirement for sequence homology.
 Rejoins double-strand breaks via direct ligation of the DNA ends without any requirement for sequence homology.")
80
Homologous recombination plays a major role in double-strand break repair in prokaryotes and single-cell eukaryotes. In mammalian cells, double-strand breaks are primarily repaired through NHEJ. In mammalian cells, the main function of homologous recombination is to repair double-strand breaks at the replication fork.

81
Homologous recombination
Many essential roles in eukaryotic organisms Crossing-over during meiosis. Transposition. Mating-type switching in yeast. Antigen-switching in trypanosomes. DNA repair.

82
Double-strand break (DSB)
Model for mammalian DNA double-strand break repair by homologous recombination Double-strand break (DSB) A DSB is induced by ionizing radiation.
 A DSB is induced by ionizing radiation.")
83
End-processing and recognition:
Recruitment of MRN (Mre11-Rad50-Nbs1) to the DSB. The 3′, 5′ exonuclease activity of Mre11 generates 3′ ssDNA tails that are recognized by Rad52.
 to the DSB. The 3′, 5′ exonuclease activity of Mre11 generates 3′ ssDNA tails that are recognized by Rad52.")
84
Strand invasion and DNA synthesis:
The 3′ tails invade homologous intact sequences. Strand exchange generates a hybrid molecule. Missing sequence information at the DSB is restored by DNA synthesis.

85
Holliday junction resolution and ligation
Branch migration Processing of the interlinked molecules. Holliday junction resolution and ligation

87
ATM activation at double-strand break (DSB) sites
ATM (ataxia telangiectasia mutated) is a serine-threonine kinase MRN complexes form a bridge between free DNA ends via Rad50. Inactive ATM is recruited to the DSBs through interaction with Nsb1.
 is a serine-threonine kinase. MRN complexes form a bridge between free DNA ends via Rad50. Inactive ATM is recruited to the DSBs through interaction with Nsb1.")
88
ATM is activated by phosphorylation.
ATM phosphorylates proteins involved in DNA repair and cell cycle control. Patients that lack ATM suffer from a syndrome called ataxia telangiectasia Extreme sensitivity to radiation Increased susceptibility to developing cancer Immunodeficiency Premature aging Neurodegenerative disorders

89
Holliday junctions Early 1960s: Robin Holliday proposed a model for general recombination based on genetic data obtained in fungi. The model has survived the test of time…

90
Heteroduplex DNA: duplex DNA formed during recombination is composed of single DNA strands originally derived from different homologs. Holliday junction: an intermediate in which the two recombining duplexes are joined covalently by single crossovers.

91
The Holliday junction is resolved into two duplexes by an enzyme complex called the resolvasome.
The resolvasome has “resolvase” activity.

92
E. coli RuvABC complex Unfolds the Holliday junction arms.
Uses ATPase activity to promote branch migration. RuvC cuts the junction symmetrically in a sequence-specific manner.

93
2004: The human Holliday junction resolvasome was purified from 50 liters of HeLa cells passed through 6 chromatographic steps. Rad51C is required for Holliday junction processing in mammalian cells. Rad51C forms a complex with the XRCC3 protein.

94
Hereditary breast cancer syndromes: mutations in BRCA1 and BRCA2
About 5-10% of all cases of breast cancer. Mutations in BRCA1 and BRCA2 “tumor suppressor genes”. Lifetime risk for breast (and ovarian) cancer: BRCA1: 50 to 87% BRCA2: 15 to 44%
 cancer: BRCA1: 50 to 87% BRCA2: 15 to 44%")
95
Nonhomologous end-joining
This double-strand break repair process can lead to mutation. Two broken ends can be ligated together regardless of whether they came from the same chromosome.

96
Nonhomologous end-joining
Frequently results in insertions or deletions at the break site. Trade-off between repair and otherwise lethal breaks in the genome.

97
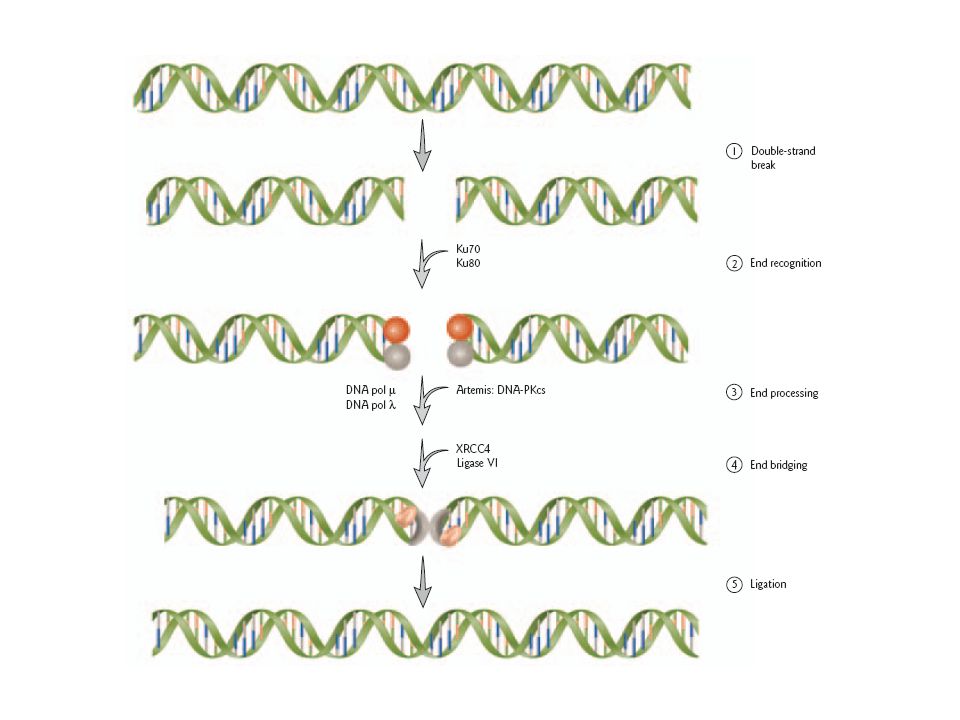
Model for mammalian DNA double-strand break repair by nonhomologous end-joining
Induced by ionizing radiation. End recognition Broken ends are recognized by heterodimers of Ku70/Ku80.

98
End processing: The endonuclease Artemis is activated by the DNA-dependent protein kinase catalytic subunit (DNA-PKCS). DNA polymerase (pol) or pol fill-in gaps and extend 3′ or 5′ overhangs.
 or pol fill-in gaps and extend 3′ or 5′ overhangs.")
99
End bridging The ligase complex XRCC4-DNA ligase IV is recruited to the damaged site and forms a bridge. Ligation The broken ends are ligated by the XRCC4-DNA ligase IV complex.

101
In vitro assays suggest that there is flexibility in the order of the three key enzymatic steps on each strand: Nucleolytic action Polymerization Ligation

Similar presentations



















 If any mismatch escapes the proof reading mechanisms it will cause distortion of the.>")











