Download presentation
Presentation is loading. Please wait.

1
DNA Metabolism: Replication, Recombination, and Repair
Chapter 28 Biochemistry by Reginald Garrett and Charles Grisham Igor Chesnokov Department of Biochemistry and Molecular Genetics Office Phone #

2
Essential Question How is this genetic information in the form of DNA replicated, how is the information rearranged, and how is its integrity maintained in the face of damage?

3
Outline How Is DNA Replicated?
What Are the Properties of DNA Polymerases? How Is DNA Replicated in Eukaryotic Cells? How Are the Ends of Chromosomes Replicated? How Are RNA Genomes Replicated? How Is the Genetic Information Shuffled by Genetic Recombination? Can DNA Be Repaired? What Is the Molecular Basis of Mutation?

4
Watson and Crick’s famous paper, in its entirety
Watson and Crick’s famous paper, in its entirety. (Reprinted with permission from Watson,J.D., and Crick,F.H.C., Molecular structure of nucleic acid, Nature 171: Copyright 1953 Macmillan Publishers Ltd.)
")
5
The Dawn of Molecular Biology
April 25, 1953 Watson and Crick: "It has not escaped our notice that the specific (base) pairing we have postulated immediately suggests a possible copying mechanism for the genetic material." The mechanism: Strand separation, followed by copying of each strand. Each separated strand acts as a template for the synthesis of a new complementary strand.
 pairing we have postulated immediately suggests a possible copying mechanism for the genetic material. The mechanism: Strand separation, followed by copying of each strand. Each separated strand acts as a template for the synthesis of a new complementary strand.")
6
How Is DNA Replicated? Replication requires unwinding of the DNA helix
DNA replication is semi-conservative DNA replication is bi-directional DNA replication is semi-discontinuous

7
Untwisting of DNA strands exposes their bases for hydrogen bonding.
Base pairing ensures that appropriate nucleotides are inserted in the correct positions as the new complementary strands are synthesized. By this mechanism, the nucleotide sequence of one strand dictates a complementary sequence in its daughter strand.

8
Three models of DNA replication prompted by Watson and Crick’s double helix structure of DNA. (a) Conservative: Each strand of the DNA duplex is replicated, and the two newly synthesized strands join to form one DNA double helix while the two parental strands remain associated with each other. The products are completely new DNA duplex and the original DNA duplex. (b) Semiconservative: The two strands separate, and each strand is copied to generate a complementary strand. Each parental strand remains associated with its newly synthesized complement, so each DNA duplex contains one parental strand and one new strand. (c) Dispersive: This model predicts that each of the four strands in the two daughter DNA duplexes contains both newly synthesized segments and segments derived from the parental strands.
 Dispersive: This model predicts that each of the four strands in the two daughter DNA duplexes contains both newly synthesized segments and segments derived from the parental strands.")
9
The Meselson and Stahl experiment demonstrating that DNA replication is semi-conservative.
On the left are densitometric traces of the DNA isolated from E. coli grown for various generation time after 15N-labeling. DNA was subjected to ultracentrifugation. Density increases from left to right. The peaks reveal the positions of the banded DNA with respect to the density of the solution. The number of generations that the E. coli cells were grown (following 14 generations of 15N density-labeling) is shown down the middle. A schematic representation interpreting the pattern expected of semiconservative replication is shown on the right side of this figure.
 is shown down the middle. A schematic representation interpreting the pattern expected of semiconservative replication is shown on the right side of this figure.")
10
DNA Replication is Bidirectional.
Bidirectional replication involves two replication forks, which move in opposite directions. By pulse labeling active replication was shown to occur at both forks. Bidirectional replication starts from a single consistent origin in the bacterial chromosome (oriC in E. coli) or multiple origins in eukaryotic chromosomes.
 or multiple origins in eukaryotic chromosomes.")
11
DNA Replication is Semi-Discontinuous.
DNA polymerase replicates both strands at once but they are in different directions & DNA replication must be 5’ to 3’ direction. One strand can be synthesized smoothly in 5’ to 3’ direction (leading strand), the other must be synthesized backwards from the replication fork (lagging strand) in discontinuous fragments (Okazaki fragments). Each fragment has own RNA primers. After primers are removed and gaps are filled, fragments are joined by DNA ligase.
, the other must be synthesized backwards from the replication fork (lagging strand) in discontinuous fragments (Okazaki fragments). Each fragment has own RNA primers. After primers are removed and gaps are filled, fragments are joined by DNA ligase.")
12
The Enzymology of DNA Replication
If Watson and Crick were right, then there should be an enzyme that makes DNA copies from a DNA template In 1957, Arthur Kornberg and colleagues demonstrated the existence of a DNA polymerase - DNA polymerase I Pol I needs all four deoxynucleotides, a template and a primer - a ss-DNA (with a free 3'-OH) that pairs with the template to form a short double-stranded region
 that pairs with the template to form a short double-stranded region.")
13
DNA Polymerase I Replication occurs 5' to 3'
Nucleotides are added at the 3'-end of the strand Pol I catalyzes about 20 cycles of polymerization before the new strand dissociates from template 20 cycles constitutes moderate "processivity" Pol I from E. coli is 928 aa (109 kD) monomer In addition to 5'-3' polymerase, it also has 3'-5' exonuclease and 5'-3' exonuclease activities
 monomer. In addition to 5 -3 polymerase, it also has 3 -5 exonuclease and 5 -3 exonuclease activities.")
15
DNA synthesis is catalyzed by DNA Polymerase.
Reaction Enzyme The reaction is driven by a large, favorable free-energy exchange, caused by release of pyrophosphate and its subsequent hydrolysis to two molecules of inorganic phosphate.

16
Nucleotides are added at the 3'-end of the strand
In addition to 5'-3' polymerase, it also has 3'-5' exonuclease and 5'-3' exonuclease activities The 3'-5' exonuclease activity serves a proofreading function! It removes incorrectly matched bases, so that the polymerase can try again Pol I, first DNA polymerase discovered, catalyzes about 20 cycles of polymerization before the new strand dissociates from template Pol I from E. coli is 928 aa (109 kD) monomer Pol III is the “real” DNA polymerase
 monomer. Pol III is the real DNA polymerase.")
17
The "real" polymerase in E. coli
DNA Polymerase III The "real" polymerase in E. coli At least 10 different subunits "Core" enzyme has three subunits - , , and Alpha subunit is polymerase Epsilon subunit is 3'-exonuclease Theta subunit is involved in holoenzyme assembly The beta subunit dimer forms a ring around DNA Enormous processivity - 5 million bases!

18
Table 28-2, p. 906

19
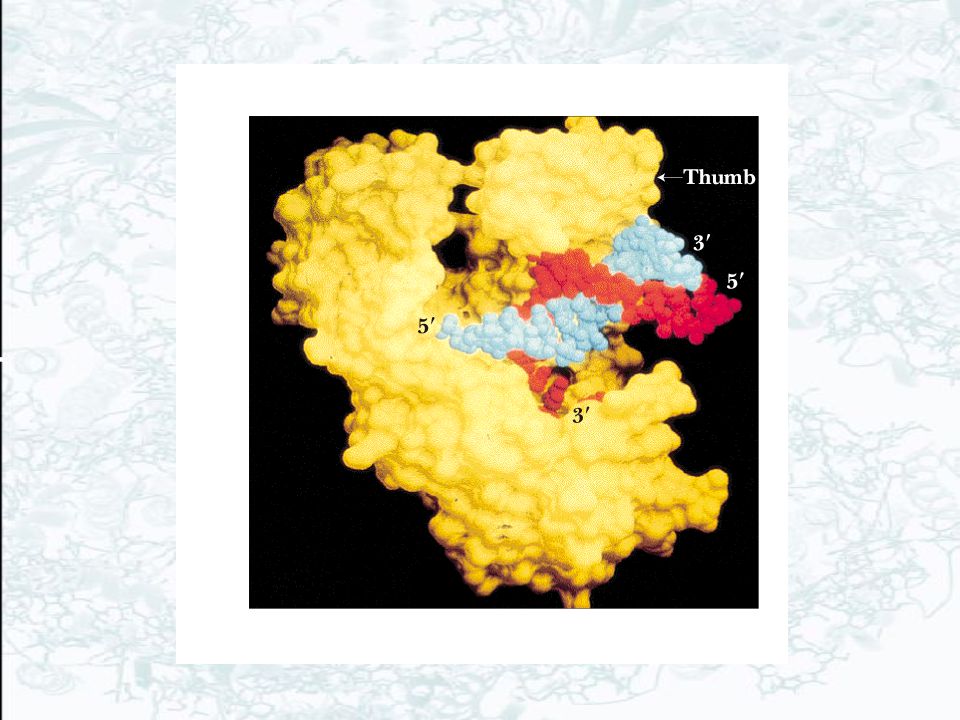
(a) Ribbon diagram of the b-subunit dimer of the DNA polymerase III holoenzyme on B-DNA viewed down the axis of the DNA. One monomer of the b-subunit dimer is colored red and the other yellow. The centrally located DNA is mostly blue. (b) Space-filling model of the b-subunit dimer of the DNA polymerase III holoenzyme on B-DNA. One monomer is shown in red, the other in yellow. The B-DNA has one strand colored white and the other blue.
 Space-filling model of the b-subunit dimer of the DNA polymerase III holoenzyme on B-DNA. One monomer is shown in red, the other in yellow. The B-DNA has one strand colored white and the other blue.")
20
General features of a replication fork.
The DNA duplex is unwound by the action of DNA gyrase and helicase, and the single strands are coated with SSB (ssDNA-binding protein). Primase periodically primes synthesis on the lagging strand (RNA primers). Each half of the dimeric replicative polymerase is a “core” polymerase bound to its template strand by a b-subunit sliding clamp. DNA polymerase I and DNA ligase act downstream on the lagging strand to remove RNA primers, replace them with DNA, and ligate the Okazaki fragments.
. Primase periodically primes synthesis on the lagging strand (RNA primers). Each half of the dimeric replicative polymerase is a core polymerase bound to its template strand by a b-subunit sliding clamp. DNA polymerase I and DNA ligase act downstream on the lagging strand to remove RNA primers, replace them with DNA, and ligate the Okazaki fragments.")
21
Table 28-3, p. 908

22
Features of Replication summary
Replication is semiconservative and bidirectional The double helix must be unwound - by helicases Supercoiling must be compensated - by DNA gyrase Replication is semi-discontinuous Leading strand is formed continuously Lagging strand is formed from Okazaki fragments

23
The proteins that initiate DNA replication in bacteria.
dnaA – major initiator protein; Primosome is composed of dnaB (helicase) and dnaG (DNA primase) dnaC is a helicase inhibitor, which is activated by the initiator proteins to load the helicase onto DNA at the replication origin and then released. This inhibitor prevents the helicase from inappropriately entering other single-stranded stretches of DNA in the bacterial genome.
 and dnaG (DNA primase) dnaC is a helicase inhibitor, which is activated by the initiator proteins to load the helicase onto DNA at the replication origin and then released. This inhibitor prevents the helicase from. inappropriately entering other single-stranded stretches of DNA in the bacterial genome.")
24
Eukaryotic DNA Replication
Like E. coli, but more complex Human cell: 6 billion base pairs of DNA to copy Multiple origins of replication: 1 per kb Several known animal DNA polymerases The cell cycle controls the timing of DNA replication

25
The phases of the cell cycle.
Two major phases of the cell cycle: S phase (DNA duplication) and M phase (chromosome segregation and cell division). Gap phases allow cells more time to grow as well as time to monitor the internal and external environment to ensure that conditions are suitable and preparations are complete for the next stage (checkpoints).
 and M phase (chromosome segregation and cell division). Gap phases allow cells more time to grow as well as time to monitor the internal and external environment to ensure that conditions are suitable and preparations are complete for the next stage (checkpoints).")
26
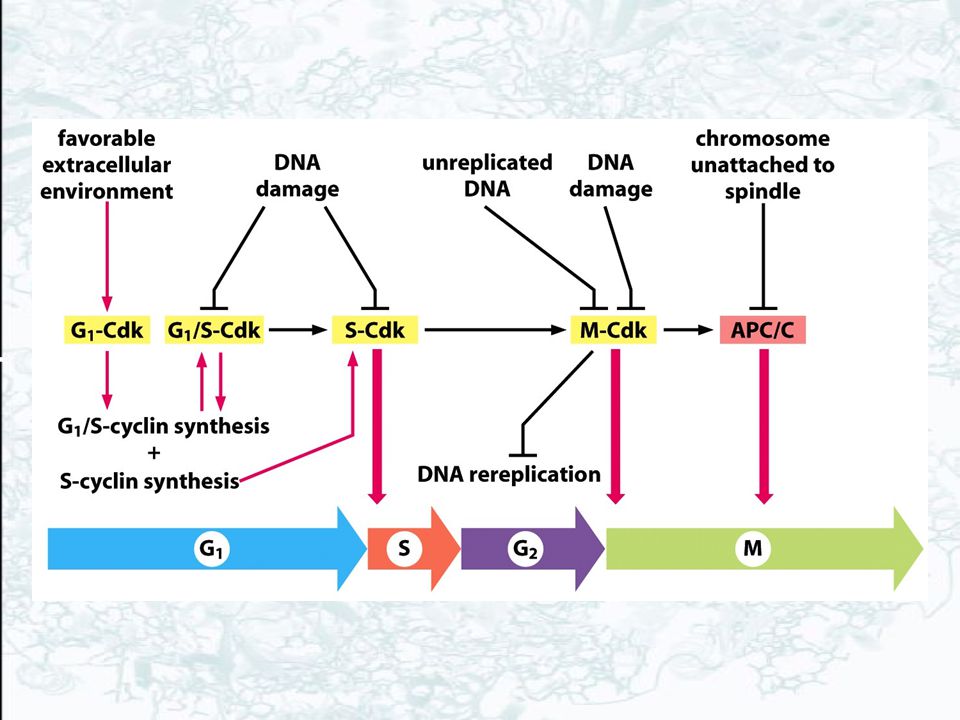
Cyclin-Cdk complexes of cell cycle control system.
Concentrations of the three major cyclin types oscillate during cell cycle. A complex of cyclin with CDK acts as a protein kinase to phosphorylate multiple protein targets to trigger specific cell cycle events. CDK activity is usually terminated by cyclin degradation. G1/S-cyclin-Cdks trigger progression through the Start checkpoint. S-cyclin-CDKs trigger DNA replication machinery. M-cyclin-CDKs trigger mitosis machinery.

28
Bacteria carry their genes on a single DNA molecule, which is usually circular.
In eukaryotes, the DNA in the nucleus is divided between a set of different chromosomes. The human genome is distributed over 24 different chromosomes. Each chromosome consists of a single, long, linear DNA molecule associated with proteins that fold and pack DNA into a more compact structure. In addition to the proteins involved in packaging the DNA, chromosomes are also associated with many proteins required for the processes of gene expression, DNA replication and DNA repair.

29
Eukaryotic chromosomes contain multiple origins of DNA replication.
Experiment – new DNA was labeled with pulse of highly radioactive thymidine. Cells are lysed and DNA are stretched out on a glass slide covered with photo emulsion. Both the rate and the direction of replication fork movement can be determined. Conclusions: 1) Replication forks travel at ~50 nucleotides per second (1/10 of the rate in bacteria) 2) Replication origins tend to activate in clusters (20-80 origins), called replication units. 3) New replication units are activated at different times during the S phase of cell cycle. 4) Within a replication unit, origins are spaced at 30, ,000 bp from one another. 5) As in bacteria, replication forks are formed in pairs and create a replication bubble as they move in opposite directions away from the origin.
 Replication forks travel at ~50 nucleotides per second (1/10 of the rate in bacteria) 2) Replication origins tend to activate in clusters (20-80 origins), called replication units. 3) New replication units are activated at different times during the S phase of cell cycle. 4) Within a replication unit, origins are spaced at 30, ,000 bp from one another. 5) As in bacteria, replication forks are formed in pairs and create a replication bubble as they move. in opposite directions away from the origin.")
30
Model for initiation of the DNA replication cycle in eukaryotes.
Origin Recognition Complex (ORC) is present at the replicators throughout the cell cycle. The pre-replication complex (pre-RC) is assembled through the sequential addition of Cdc6, MCM and other proteins during a window of opportunity defined by the state of the cyclin-CDKs. After initiation, a post-RC state is established.
 is present at the replicators throughout the cell cycle. The pre-replication complex (pre-RC) is assembled through the sequential addition of Cdc6, MCM and other proteins during a window of opportunity defined by the state of the cyclin-CDKs. After initiation, a post-RC state is established.")
31
The Initiation of DNA Replication in Eukaryotic Cells.
Binding of the pre-RC to origins of replication is followed by loading of MCM hexameric helicases, phosphorylation reactions mediated by S-CDK and Cdc7-Dbf4, and binding of the complex.

32
The Initiation of DNA Replication in Eukaryotic Cells.
Phosphorylation of Sld2 and Sld3 leads to the recruitment of DNA polymerase to the replication origins. The two diverging MCM complexes serve as helicases, providing single-strand templates.

33
Possible Mechanism of Replicative Helicase Loading.
Individual subunits recruited to the origin and assembled around DNA molecule. The helicase is recruited as a hexamer, loading onto DNA assisted by other proteins. Hypothetical mechanism for the opening of a MCM hexamer by Orc1, Orc4, Orc5 and Cdc6, four other AAA+ ATPases present at the origins at the time of MCM loading.

34
Proteins that perform analogous functions at replication forks.
E.Coli/Phage l Human Helicase Primase Polymerase Proofreading Exonuclease Sliding clamp Clamp loader Single strand DNA binding protein DnaB DnaG a-subunit of DNA pol III e-subunit of b-subunit g-subunit SSB MCM proteins T antigen (SV40) Primase subunit of Pol a primase Pol d and e Part of polymerase Subunit of pol d PCNA RF-C RP-A
 Primase subunit of. Pol a primase. Pol d and e. Part of polymerase. Subunit of pol d. PCNA. RF-C. RP-A.")
35
Table 28-4, p. 912

36
A mammalian replication fork.
DNA polymerase e There are more protein components in eukaryotic replication machines, even though the basic functions are the same (RP-A – 3 subunits, bacterial SSB – 1 subunit). Two different DNA polymerases on the lagging strand. The mammalian DNA primase is a subunit of DNA pol a. In bacteria primase associates with DNA helicase in primosome. Pol a with its associated primase begins chains with RNA, extends them with DNA and then hands chains over to Pol d, which elongates them.
. Two different DNA polymerases on the lagging strand. The mammalian DNA primase is a subunit of DNA pol a. In bacteria primase associates with DNA helicase in primosome. Pol a with its associated primase begins chains with RNA, extends them with DNA and then hands chains over to Pol d, which elongates them.")
37
How Are the Ends of Chromosomes Replicated?
Telomeres, the structures at the ends of eukaryotic chromosomes, consist of 5-8 bp tandemly repeated G-rich nucleotide sequences Telomeres are 1-12 kb long Telomeres are replicated by an RNA-dependent DNA polymerase called telomerase

38
Telomere replication. During replication of the lagging strand, short RNA primers are added (pink) and then extended by DNA polymerase. When the RNA prirmer at the 5¢-end of each strand is removed, there is no nucleotide sequence to read in the next round of DNA replication. The result is a gap (primer gap) at the 5¢-end of each strand (only one end of the chromosome is shown in this figure). Synthesis of telomeric DNA by telomerase extends the 5¢-ends of DNA strands, allowing the strands to be copied by normal DNA replication. Asterisks indicate sequences at the 3¢-end that cannot be copied by conventional DNA replication.
 and then extended by DNA polymerase. When the RNA prirmer at the 5¢-end of each strand is removed, there is no nucleotide sequence to read in the next round of DNA replication. The result is a gap (primer gap) at the 5¢-end of each strand (only one end of the chromosome is shown in this figure). Synthesis of telomeric DNA by telomerase extends the 5¢-ends of DNA strands, allowing the strands to be copied by normal DNA replication. Asterisks indicate sequences at the 3¢-end that cannot be copied by conventional DNA replication.")
39
Telomerase replicates the ends of chromosomes.
Telomerase is a protein-RNA complex that uses an internal RNA template and reverse transcriptase activity for synthesizing a repeating G-rich telomere DNA sequence - (TTAGGG)n in mammals. Replication of the lagging strand can then be completed by using these new extensions as a template by DNA polymerase.
n in mammals. Replication of the lagging strand can then be completed by using these new extensions as a template by DNA polymerase.")
40
A mammalian replication fork.
DNA Replication animation from cshl.org -

Similar presentations



 Two strands of DNA run antiparallel.>")















