Download presentation
Presentation is loading. Please wait.

1
Chapter 14 Mechanisms of Enzyme Action Biochemistry by
Reginald Garrett and Charles Grisham

2
Essential Question Although the catalytic properties of enzymes may seem almost magical, it is simply chemistry– the breaking and making of bonds– that give enzymes their prowess What are the universal chemical principles that influence the mechanisms of these and other enzymes How may we understand the many other cases, in light of the knowledge gained from these examples?

3
Outline of Chapter 14 What Role Does Transition-State Stabilization Play in Enzyme Catalysis? What Are the Magnitudes of Enzyme-Induced Rate Accelerations? Why Is the Binding Energy of ES Crucial to Catalysis? What Roles Do Entropy Loss and Destabilization of the ES Complex Play? How Tightly Do Transition-State Analogs Bind to the Active Site? What Are the Mechanisms of Catalysis? What Can Be Learned from Typical Enzyme Mechanisms?

4
14.1 – What Role Does Transition-State Stabilization Play in Enzyme Catalysis?
H-O-H + Cl H-O H Cl HO- + HCl Reactant Transition state Products Transition state (10-13sec) Intermediate (10-13~10-3sec) d- d-
 Intermediate (10-13~10-3sec) d- d-")
5
Figure 14.1 Enzymes catalyze reactions by lowering the activation energy. Here the free energy of activation for (a) the uncatalyzed reaction, DGu‡, is larger than that for (b) the enzyme-catalyzed reaction, DGe‡.
 the uncatalyzed reaction, DGu‡, is larger than that for (b) the enzyme-catalyzed reaction, DGe‡..")
6
Reaction rate acceleration by an enzyme means that the energy barrier between ES and EX‡ must be smaller than the barrier between S and X‡ (DGe ‡ <DGu‡) This means that the enzyme must stabilize the EX‡ transition state more than it stabilizes ES Enzymes bind the transition state structure more tightly than the substrate

7
14.2 – What Are the Magnitudes of Enzyme-Induced Rated Accelerations?

8
14.2 – What Are the Magnitudes of Enzyme-Induced Rated Accelerations?
Mechanisms of catalysis: Entropy loss in ES formation Destabilization of ES Covalent catalysis General acid-base catalysis Metal ion catalysis Proximity and orientation

9
14.3 – Why Is the Binding Energy of ES Crucial to Catalysis?
The favorable interactions between the substrate and amino acid residues on the enzyme account for the intrinsic binding energy, DGb The intrinsic binding energy ensures the favorable formation of the ES complex, but not too favorable!

10
intrinsic binding energy
Figure 14.2 The intrinsic binding energy of the enzyme-substrate (ES) complex (DGb ) is compensated to some extent by entropy loss due to the binding of E and S (TDS) and by destabilization of ES (DGd) by strain, distortion, desolvation , and similar effects. If DGb were not compensated by TDS and DGd, the formation of ES would follow the dashed line.
 complex (DGb ) is compensated to some extent by entropy loss due to the binding of E and S (TDS) and by destabilization of ES (DGd) by strain, distortion, desolvation , and similar effects. If DGb were not compensated by TDS and DGd, the formation of ES would follow the dashed line.")
11
Figure 14.3 (a) Catalysis does not occur if the ES complex and the transition state for the reaction are stabilized to equal extents. (b) Catalysis will occur if the transition state is stabilized to a greater extent than the ES complex (right). Entropy loss and destabilization of the ES complex DGd ensure that this will be the case.
 Catalysis will occur if the transition state is stabilized to a greater extent than the ES complex (right). Entropy loss and destabilization of the ES complex DGd ensure that this will be the case..")
12
14.3 – Why Is the Binding Energy of ES Crucial to Catalysis?
If uncompensated, it makes the activation energy for the enzyme-catalyzed reaction unnecessarily large and wastes some of the catalytic power of the enzyme

13
Raising the energy of ES raises the rate
14.4 – What Roles Do Entropy Loss and Destabilization of the ES Complex Play? Raising the energy of ES raises the rate For a given energy of EX‡, raising the energy of ES will increase the catalyzed rate This is accomplished by loss of entropy due to formation of ES destabilization of ES by structural strain & distortion desolvation electrostatic effects

14
Figure 14. 4 Formation of the ES complex results in a loss of entropy
Figure Formation of the ES complex results in a loss of entropy. Prior to binding, E and S are free to undergo translational and rotational motion. By comparison, the ES complex is a more highly ordered, low-entropy complex.

15
Figure 14.5 Substrates typically lose waters of hydration in the formation of the ES complex. Desolvation raises the energy of the ES complex, making it more reactive. Figure 14.6 Electrostatic destabilization of a substrate may arise from juxtaposition of like charges in the active site. If such charge repulsion is relieved in the course of the reaction, electrostatic destabilization can result in a rate increase.

16
14.5 – How Tightly Do Transition-State Analogs Bind to the Active Site?
Transition state is exists only for about sec, less than the time required for a bond vibration The nature of the elusive transition state can be explored using transition state analogs Transition state analogs are stable molecules, chemically and structurally similar to the transition state Transition-state analogs are only approximations of the transition state itself and will never bind as tightly as would be expected for the true transition state

17
Transition-State Analogs
Figure 14.7 The proline racemase reaction. Pyrrole-2-carboxylate and D -1-pyrroline-2-carboxylate mimic the planar transition state of the reaction.

18
Figure 14.8 (a) Phosphoglycolohydroxamate is an analog of the enediolate transition state of the yeast aldolase reaction. (b) Purine riboside, a potent inhibitor of the calf intestinal adenosine deaminase reaction, binds to adenosine deaminase as the 1,6-hydrate. The hydrated form of purine riboside is an analog of the proposed transition state for the reaction.
 Purine riboside, a potent inhibitor of the calf intestinal adenosine deaminase reaction, binds to adenosine deaminase as the 1,6-hydrate. The hydrated form of purine riboside is an analog of the proposed transition state for the reaction..")
19
14.6 – What Are the Mechanisms of Catalysis?
Covalent catalysis Some enzyme reactions derive much of their rate acceleration from the formation of covalent bonds between enzyme and substrate BX + Y BY + X BX + Enz E:B + X + Y Enz + BY Most enzymes that carry out covalent catalysis have ping-pong kinetic mechanisms

20
14.6 – What Are the Mechanisms of Catalysis?
Covalent catalysis The side chains of amino acids in proteins offer a variety of nucleophilic centers for catalysis, including amines, carboxylate, aryl and alkyl hydroxyls, imidazoles, and thiol groups These groups are readily attack electrophilic centers of substrates, forming covalently bonded enzyme-substrate intermediate

21
Figure 14.9 Examples of covalent bond formation between enzyme and substrate. In each case, a nucleophilic center (X:) on an enzyme attacks an electrophilic center on a substrate.
 on an enzyme attacks an electrophilic center on a substrate..")
23
Glyceraldehyde-3-P + NAD+ + Pi 1,3-bisphosphoglycerate + NADH + H+
Figure 14.10 Formation of a covalent intermediate in the glyceraldehyde-3-phosphate dehydrogenase reaction. Nucleophilic attack by a cysteine —SH group forms a covalent acylcysteine intermediate. Following hydride transfer to NAD+, nucleophilic attack by phosphate yields the product, 1,3-bisphosphoglycerate.

24
14.6 – What Are the Mechanisms of Catalysis?
General acid-base catalysis Specific acid-base catalysis involves H+ or OH- that diffuses into the catalytic center General acid-base catalysis involves acids and bases other than H+ and OH- These other acids and bases facilitate transfer of H+ in the transition state

25
Figure Specific and general acid - base catalysis of simple reactions in solution may be distinguished by determining the dependence of observed reaction rate constants (kobs) on pH and buffer concentration. (a) In specific acid-base catalysis, H+ or OH- concentration affects the reaction rate, kobs is pH-dependent, but buffers (which accept or donate H+/OH-) have no effect. (b) In general acid - base catalysis, in which an ionizable buffer may donate or accept a proton in the transition state, kobs is dependent on buffer concentration.
 In specific acid-base catalysis, H+ or OH- concentration affects the reaction rate, kobs is pH-dependent, but buffers (which accept or donate H+/OH-) have no effect. (b) In general acid - base catalysis, in which an ionizable buffer may donate or accept a proton in the transition state, kobs is dependent on buffer concentration..")
26
Figure 14.12 Catalysis of p-nitrophenylacetate hydrolysis by imidazole—an example of general base catalysis. Proton transfer to imidazole in the transition state facilitates hydroxyl attack on the substrate carbonyl carbon.

27
14.6 – What Are the Mechanisms of Catalysis?
Low-barrier hydrogen bond (LBHB) The typical strength of a hydrogen bond is 10 to 30 kJ/mol 0.25 nm (LBHB) 0.5 order 0.1 nm nm 1 order
 The typical strength of a hydrogen bond is 10 to 30 kJ/mol nm (LBHB) 0.5 order. 0.1 nm 0.18nm. 1 order")
28
Low-Barrier Hydrogen Bonds
As distance between heteroatoms becomes smaller, H bonds become stronger Stabilization energies of LBHB may approach 60 kJ/mol in solution pKa values of the two electronegative atoms must be similar Energy released in forming an LBHB can assist catalysis

29
14.6 – What Are the Mechanisms of Catalysis?
Metal ion catalysis Many enzymes require metal ions for maximal activity (metalloenzymes) Stabilizing the increased electron density or negative charge Provide a powerful nucleophile at neutral pH M NucH M2+(NucH) M2+(NucH) + H+
 Stabilizing the increased electron density or negative charge. Provide a powerful nucleophile at neutral pH. M2+ + NucH M2+(NucH) M2+(NucH) + H+")
30
14.6 – What Are the Mechanisms of Catalysis?
Proximity Chemical reactions go faster when the reactants are in proximity, that is, near each other Proximity and orientation play a role in enzyme catalysis, but there is a problem with each of the aforementioned comparisons

31
Figure 14. 15 An example of proximity effects in catalysis
Figure An example of proximity effects in catalysis. (a) The imidazole-catalyzed hydrolysis of p-nitrophenylacetate is slow, but (b) the corresponding intramolecular reaction is 24-fold faster (assuming [imidazole] = 1 M in [a]).
 The imidazole-catalyzed hydrolysis of p-nitrophenylacetate is slow, but (b) the corresponding intramolecular reaction is 24-fold faster (assuming [imidazole] = 1 M in [a]).")
32
Figure Orientation effects in intramolecular reactions can be dramatic. Steric crowding by methyl groups provides a rate acceleration of 2.5 X 1011 for the lower reaction compared to the upper reaction. (Adapted from Milstien,S., and Cohen, L.A., Stereopopulation control I. Rate enhancements in the lactonization of o-hydroxyhydrocinnamic acid. Journal of the American Chemical Society 94: )
.")
33
14.7 – What Can Be Learned from Typical Enzyme Mechanisms?
Serine proteases and aspartic proteases are good examples Knowledge of the tertiary structure of an enzyme is important Enzymes are the catalytic machines that sustain life

34
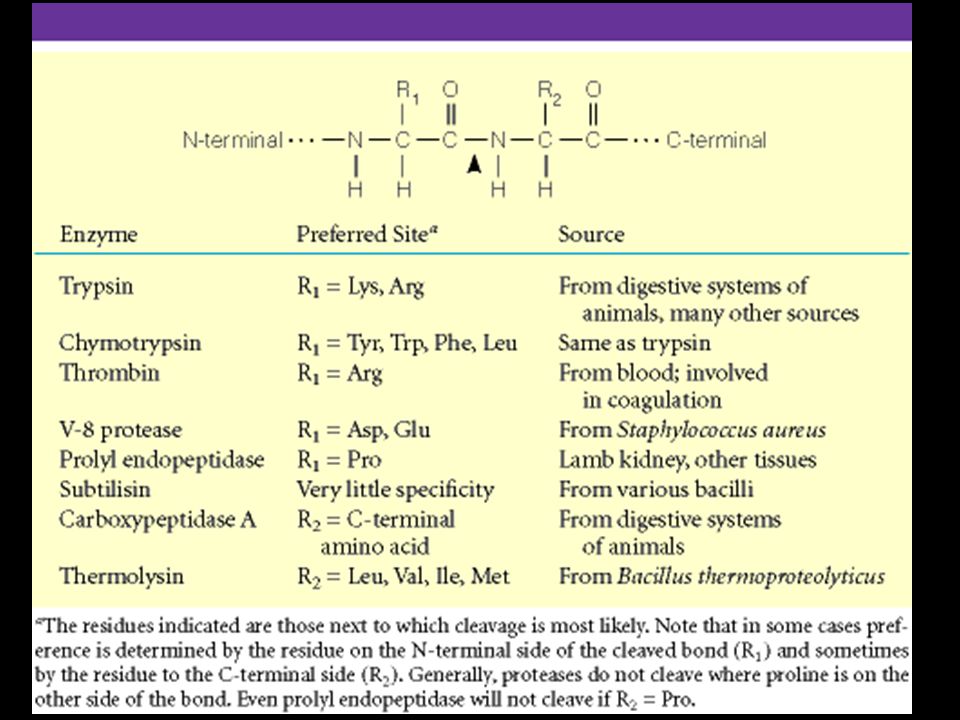
Trypsin, chymotrypsin, elastase, thrombin, subtilisin, plasmin, TPA
The Serine Proteases Trypsin, chymotrypsin, elastase, thrombin, subtilisin, plasmin, TPA Serine proteases are a class of proteolytic enzymes whose catalytic mechanism is based on an active-site serine residue Ser is part of a "catalytic triad" of Ser, His, Asp Serine proteases are homologous, but locations of the three crucial residues differ somewhat Enzymologists agree, however, to number them always as His-57, Asp-102, Ser-195

35
Figure Comparison of the amino acid sequences of chymotrypsinogen, trypsinogen, and elastase. Each circle represents one amino acid. Numbering is based on the sequence of chymotrypsinogen. Filled circles indicate residues that are identical in all three proteins. Disulfide bonds are indicated in yellow. The positions of the three catalytically important active-site residues (His57, Asp102, and Ser195) are indicated.
 are indicated..")
37
Serine Protease Mechanism
A mixture of covalent and general acid-base catalysis Asp-102 functions only to orient His-57 His-57 acts as a general acid and base Ser-195 forms a covalent bond with peptide to be cleaved Covalent bond formation turns a trigonal C into a tetrahedral C The tetrahedral oxyanion intermediate is stabilized by N-Hs of Gly-193 and Ser-195

38
Figure Structure of chymotrypsin (white) in a complex with eglin C (blue ribbon structure), a target protein. The residues of the catalytic triad (His57, Asp102, and Ser195) are highlighted. His57 (blue) is flanked above by Asp102 (red) and on the right by Ser195 (yellow). The catalytic site is filled by a peptide segment of eglin. Note how close Ser195 is to the peptide that would be cleaved in a chymotrypsin reaction. Figure The catalytic triad of chymotrypsin .

39
Figure The substrate-binding pockets of trypsin, chymotrypsin, and elastase. (Illustration: Irving Geis. Rights owned by Howard Hughes Medical Institute. Not to be reproduced without permission. )
.")
40
Serine Protease Mechanism
Kinetics The mechanism is based on studies of the hydrolysis of artificial substrates– simple organic ester Figure Artificial substrates used in studies of the mechanism of chymotrypsin .

41
Multistep mechanism Figure Burst kinetics observed in the chymotrypsin reaction. A burst of nitrophenolate production is followed by a slower, steady-state release. After an initial lag period, acetate release is also observed. This kinetic pattern is consistent with rapid formation of an acyl-enzyme intermediate (and the burst of nitrophenolate ). The slower, steady-state release of products corresponds to rate-limiting breakdown of the acyl-enzyme intermediate.
. The slower, steady-state release of products corresponds to rate-limiting breakdown of the acyl-enzyme intermediate.")
42
Serine Protease Mechanism
Kinetics In the chymotrypsin mechanism, the nitrophenylacetate combines with the enzyme to form an ES complex Followed by a rapid second step in which an acyl-enzyme intermediate is formed, with the acetyl group covalently bonded to the very reactive Ser-195

43
Figure 14.23 Rapid formation of the acyl-enzyme intermediate is followed by slower product release.

44
Multistep mechanism Figure Burst kinetics observed in the chymotrypsin reaction. A burst of nitrophenolate production is followed by a slower, steady-state release. After an initial lag period, acetate release is also observed. This kinetic pattern is consistent with rapid formation of an acyl-enzyme intermediate (and the burst of nitrophenolate ). The slower, steady-state release of products corresponds to rate-limiting breakdown of the acyl-enzyme intermediate.
. The slower, steady-state release of products corresponds to rate-limiting breakdown of the acyl-enzyme intermediate.")
45
Figure Diisopropylfluorophosphate (DIFP) reacts with active-site serine residues of serine proteases (and esterases), causing permanent inactivation.
 reacts with active-site serine residues of serine proteases (and esterases), causing permanent inactivation.")
46
Figure 14. 25 A detailed mechanism for the chymotrypsin reaction
Figure A detailed mechanism for the chymotrypsin reaction. Note the low-barrier hydrogen bond (LBHB) in (c) and (g). ES complex Tetrahedral oxyanion transition state Acyl-enzyme intermediate Acyl-enzyme-H2O intermediate Tetrahedral oxyanion transition state
 in (c) and (g). ES complex. Tetrahedral oxyanion transition state. Acyl-enzyme intermediate. Acyl-enzyme-H2O intermediate. Tetrahedral oxyanion transition state.")
47
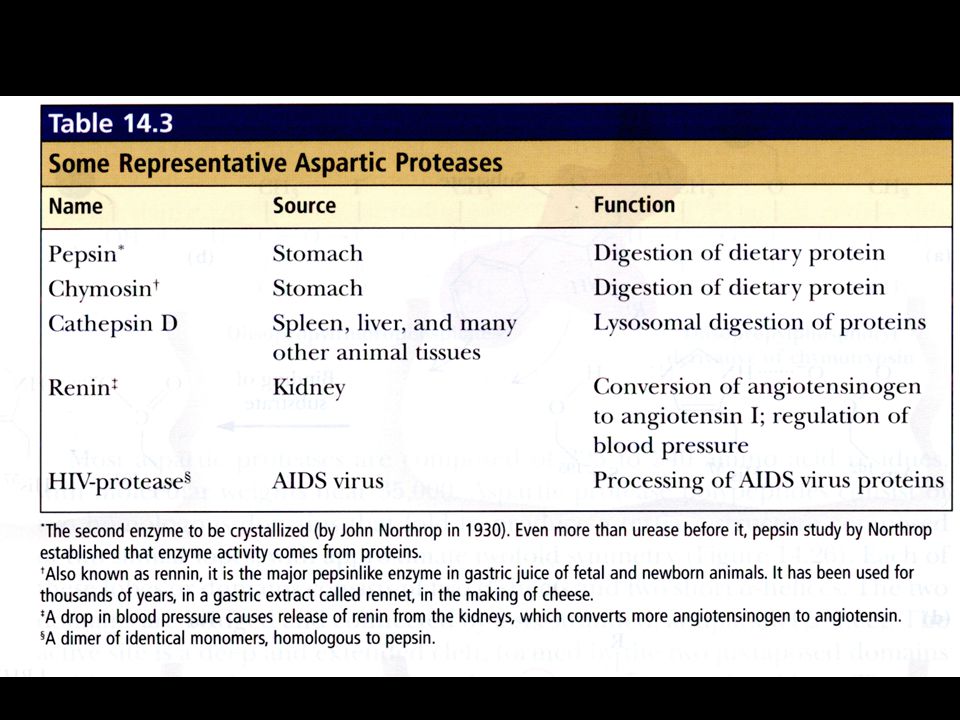
The Aspartic Proteases
Pepsin, chymosin, cathepsin D, renin and HIV-1 protease All involve two Asp residues at the active site and two Asps work together as general acid-base catalysts Active at acidic pH Most aspartic proteases have a tertiary structure consisting of two lobes (N-terminal and C-terminal) with approximate two-fold symmetry (fig 14.26) HIV-1 protease is a homodimer
 with approximate two-fold symmetry (fig 14.26) HIV-1 protease is a homodimer.")
49
Figure Structures of (a) HIV-1 protease, a dimer, and (b) pepsin (a monomer). Pepsin’s N-terminal half is shown in red; C-terminal half is shown in blue.

50
Figure Acyl-enzyme and amino-enzyme intermediates originally proposed for aspartic proteases were modeled after the acyl-enzyme intermediate of the serine proteases.

51
Aspartic Protease Mechanism
pH dependence (fig 14.28) The aspartate carboxyl groups functioned alternately as general acid and general base Deprotonated Asp acts as general base, accepting a proton from HOH, forming OH- in the transition state Other Asp (general acid) donates a proton, facilitating formation of tetrahedral intermediate
 The aspartate carboxyl groups functioned alternately as general acid and general base. Deprotonated Asp acts as general base, accepting a proton from HOH, forming OH- in the transition state. Other Asp (general acid) donates a proton, facilitating formation of tetrahedral intermediate.")
52
Figure 14. 28 pH-rate profiles for (a) pepsin and (b) HIV protease
Figure pH-rate profiles for (a) pepsin and (b) HIV protease. (Adapted fro Denburg,J., et at., The effect of PH on the rates of hydrolysis of three acylated dipeptiedes by pepsin.
 pepsin and (b) HIV protease. (Adapted fro Denburg,J., et at., The effect of PH on the rates of hydrolysis of three acylated dipeptiedes by pepsin.")
53
Figure 14. 29 A mechanism for the aspartic proteases
Figure A mechanism for the aspartic proteases. The letter titles describe the state as follows: E represents the enzyme form with a low-barrier hydrogen bond between the catalytic aspartates, F represents the enzyme form with one aspartate protonated and the other sharing in the conventional hydrogen bond, S represents bound substrate, T represents a tetrahedral amide hydrate intermediate, P represents bound carboxyl product, and Q represents bound amine product. This mechanism is based in part on a mechanism proposed by Dexter Northrop, a distant relative of John Northrop, who had first crystallized pepsin in The mechanism is also based on data of Thomas Meek.

54
A novel aspartic protease
HIV-1 Protease A novel aspartic protease HIV-1 protease cleaves the polyprotein products of the HIV genome, producing several proteins necessary for viral growth and cellular infection This is a remarkable imitation of mammalian aspartic proteases HIV-1 protease is a homodimer - more genetically economical for the virus Active site is two-fold symmetric Two aspartate residues, Asp-25 and Asp-25’

55
Figure HIV mRNA provides the genetic information for synthesis of a polyprotein . Proteolytic cleavage of this polyprotein by HIV protease produces the individual proteins required for viral growth and cellular infection.

56
Figure (left) HIV-1 protease complexed with the inhibitor Crixivan (red) made by Merck. The flaps (residues from each subunit) covering the active site are shown in green and the active site aspartate residues involved in catalysis are shown in white. (right) The close-up of the active site shows the interaction of Crixivan with the carboxyl groups of the essential aspartate residues.
 covering the active site are shown in green and the active site aspartate residues involved in catalysis are shown in white. (right) The close-up of the active site shows the interaction of Crixivan with the carboxyl groups of the essential aspartate residues..")
57
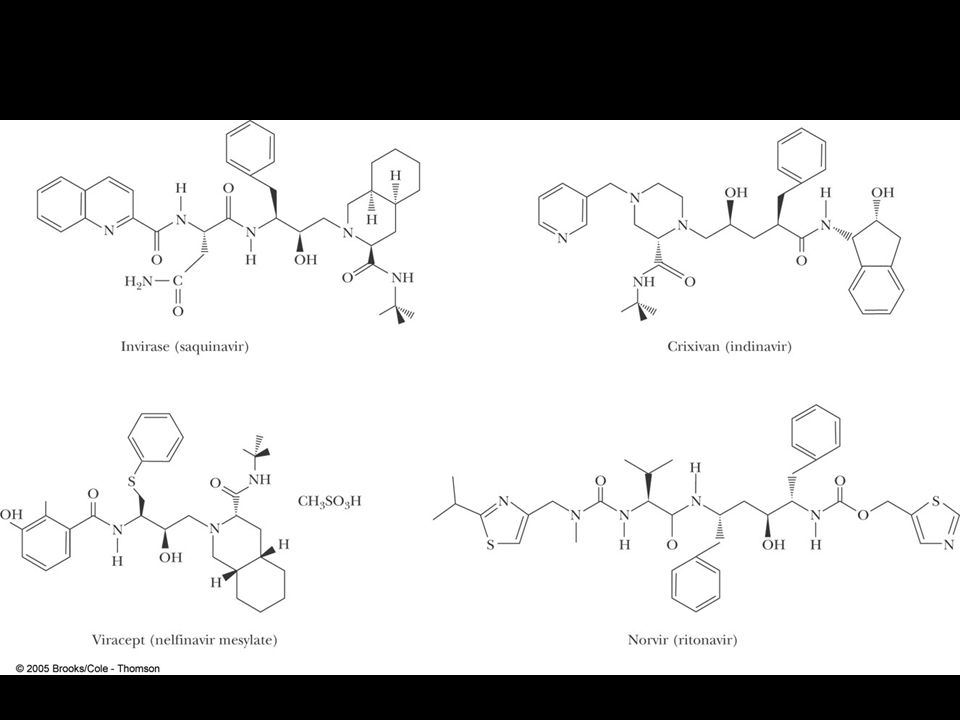
Protease inhibitors as AIDS drugs
Therapy for HIV? Protease inhibitors as AIDS drugs If the HIV-1 protease can be selectively inhibited, then new HIV particles cannot form Several novel protease inhibitors are currently marketed as AIDS drugs Many such inhibitors work in a culture dish However, a successful drug must be able to kill the virus in a human subject without blocking other essential proteases in the body

59
Lysozyme Lysozyme hydrolyzes polysaccharide chains and ruptures certain bacterial cells by breaking down the cell wall Hen egg white enzyme has 129 residues with four disulfide bonds (8 cysteine residues) Lysozyme hydrolyzes the glycosidic bond between C-1 of NAM and C-4 of NAG
 Lysozyme hydrolyzes the glycosidic bond between C-1 of NAM and C-4 of NAG.")
60
Figure 14.33 The lysozyme reaction.

61
Figure 14. 34 The structure of lysozyme
Figure The structure of lysozyme. Glu35 and Asp52 are shown in white.

62
Substrate Analog Studies
Natural substrates are not stable in the active site for structural studies But analogs can be used - like (NAG)3 Fitting a NAG of (NAG)6 into the D site requires a distortion of the sugar This argues for stabilization of a transition state via destabilization (distortion and strain) of the substrate
3. Fitting a NAG of (NAG)6 into the D site requires a distortion of the sugar. This argues for stabilization of a transition state via destabilization (distortion and strain) of the substrate.")
63
Figure 14.35 (NAG)3, a substrate analog, forms stable complexes with lysozyme .
Figure The lysozyme -enzyme-substrate complex. (Photo courtesy of John Rupley , University of Arizona )
")
65
Figure Enzyme-substrate interactions at the six sugar residue-binding subsites of the lysozyme active site. (Illustration: Irving Geis. Rights owned by Howard Hughes Medical Institute. Not to be reproduced without permission. )
.")
66
The Lysozyme Mechanism
Studies with 18O-enriched water show that the C1-O bond is cleaved on the substrate between the D and E sites This incorporates 18O into C1 position of the sugar at the D site, not into the oxygen at C4 at the E site Glu35 acts as a general acid Distortion of the sugar ring at the D site Asp52 stabilizes a carbocation intermediate

67
Figure The C1-O bond, not the O-C4 bond, is cleaved in the lysozyme reaction. 18O from H218O is thus incorporated at the C1 position.

68
Figure 14. 39 Two possible mechanisms for the lysozyme reaction
Figure Two possible mechanisms for the lysozyme reaction. In Path A, the intermediate is a noncovalent oxocarbenium ion (carbocation). Path B depends upon a covalent intermediate involving Asp52 and the C-1 oxygen of the cleaved glycosidic bond.
. Path B depends upon a covalent intermediate involving Asp52 and the C-1 oxygen of the cleaved glycosidic bond.")
69
Figure 14. 40 Mass spectra of lysozyme complexes
Figure Mass spectra of lysozyme complexes. (a) Wild-type hen egg white lysozyme (HEWL). (b) Mutant lysozyme with Glu35 replaced with glutamine [HEWL(E35Q)], incubated with chitobiosyl fluoride (NAG2F). (c) Wild-type HEWL, incubated with 2-acetamido-2-deoxy-b-D-glucopyranosyl-(1®4)-2-deoxy-2-fluoro-b-D-glucopyroanosyl fluoride (NAG2FGlcF). (d) HEWL(E35Q), incubated with NAG2FGlcF. Structures of the species corresponding to each peak observed in the amss spectra are shown to the right, with their expected relatvie molecular mass. (From Vocadlo,D.J.,et al., Catalysis by hen egg-white lysozyme proceeds via a covalent intermediate. Nature 412: )
 Wild-type hen egg white lysozyme (HEWL). (b) Mutant lysozyme with Glu35 replaced with glutamine [HEWL(E35Q)], incubated with chitobiosyl fluoride (NAG2F). (c) Wild-type HEWL, incubated with 2-acetamido-2-deoxy-b-D-glucopyranosyl-(1®4)-2-deoxy-2-fluoro-b-D-glucopyroanosyl fluoride (NAG2FGlcF). (d) HEWL(E35Q), incubated with NAG2FGlcF. Structures of the species corresponding to each peak observed in the amss spectra are shown to the right, with their expected relatvie molecular mass. (From Vocadlo,D.J.,et al., Catalysis by hen egg-white lysozyme proceeds via a covalent intermediate. Nature 412: )")
70
Figure Stereo view of the ovalent NAG2FGlcF intermediate of the lysozyme reaction. (From Vacadlo,D.J.,et al., Catalysis by hen egg-white lysozyme proceeds via a covalent intermediate. Nature 412: )
.")
Similar presentations