Download presentation
Presentation is loading. Please wait.

1
Молекулярная биология Транскрипция
ДВАДЦАТЬ ШЕСТАЯ ЛЕТНЯЯ МНОГОПРЕДМЕТНАЯ ШКОЛА с. Вишкиль. 3 – 28 июля 2010 года Биологическое отделение Молекулярная биология Транскрипция Вишкиль, 2010

2
Определение: Транскрипция генов – процесс биосинтеза молекул РНК, комплементарных одной из цепей матричной ДНК, сопровождаемый полимеризацией четырех рибонуклеозидтрифосфатов (ATP, GTP, CTP и UTP) с образованием 3’– 5’-фосфодиэфирных связей и освобождением неорганического пирофосфата.
 с образованием 3’– 5’-фосфодиэфирных связей и освобождением неорганического пирофосфата.")
3
транскрипция трансляция ДНК РНК белок репликация

4
Сравнение транскрипции и репликации
Инициация в специфических участках – промоторах Инициация синтеза РНК с мононуклеотидов, без затравки Использование рибонуклеозидтрифосфатов Копирование одной из цепей ДНК (матричная цепь) Направление синтеза от 5’- к 3’-концу РНК РНК-полимераза нематричная ДНК 5’ 3’ 3’ 5’ 3’ РНК матричная ДНК 5’
 Направление синтеза от 5’- к 3’-концу РНК. РНК-полимераза. нематричная ДНК. 5’ 3’ 3’ 5’ 3’ РНК. матричная ДНК. 5’")
5
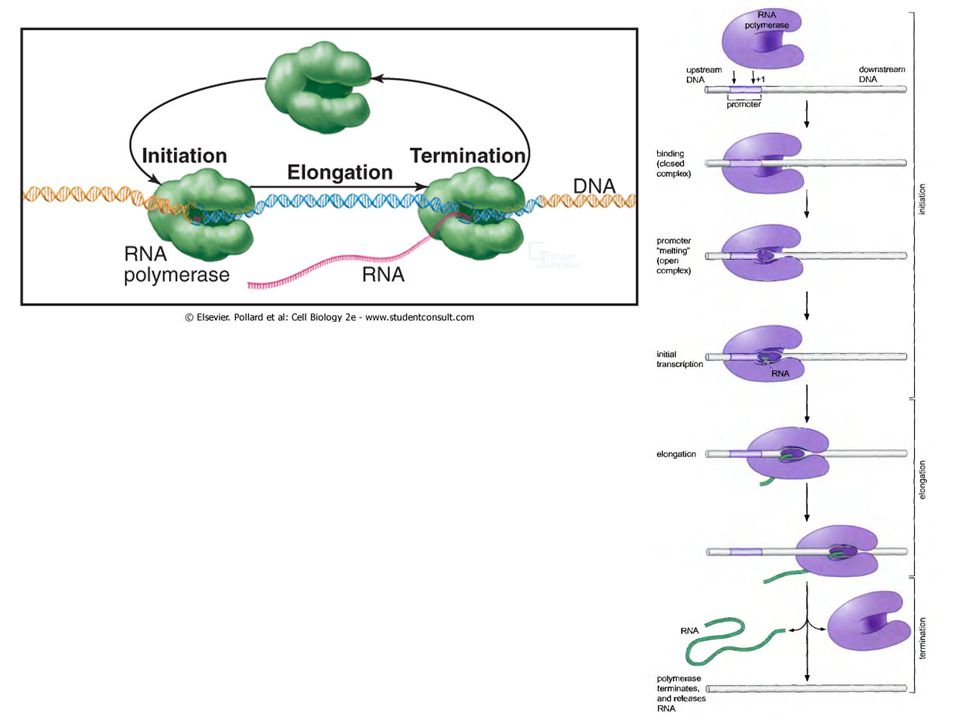
Транскрипционный цикл
промотор Инициация (промотор) Терминация (терминатор) Для узнавания промоторов нужны дополнительные белки Основная стадия регуляции Элонгация
 Терминация. (терминатор) Для узнавания промоторов нужны дополнительные белки. Основная стадия регуляции. Элонгация.")
6
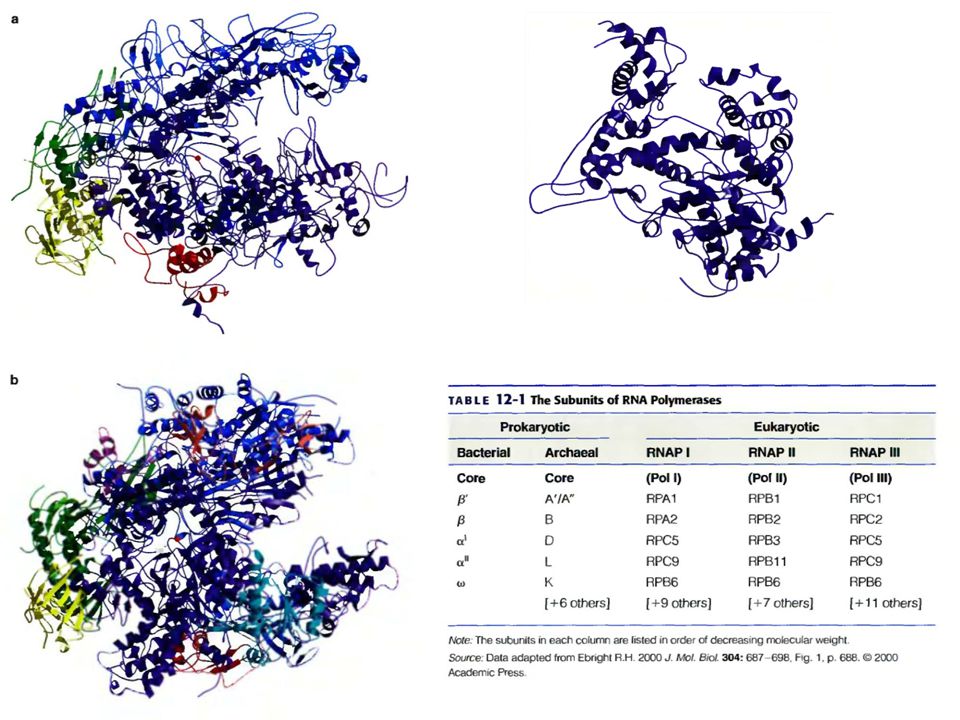
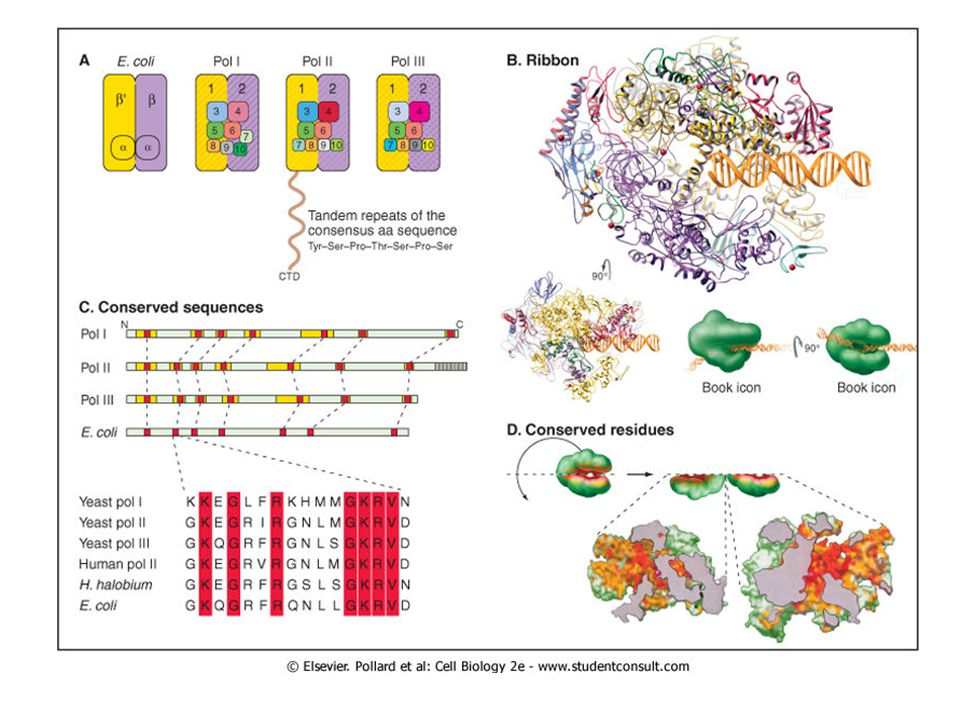
Строение РНК-полимеразы:
Полный набор субъединиц, входящих в состав РНК-полимеразы, называют холоферментом. У E.coli он представлен агрегатом, весом 465 кДа, в который входят 4 различные субъединицы:α, β, β’, δ.

7
Строение РНК-полимеразы:
Β и β’ субъединицы вместе формируют каталитический центр, они гомологичны по аминокислотной последовательности большой субъединице эукариотических РНК-полимераз, что доказывает общность происхождения всех РНК-полимераз. Мутации в генах, кодирующих данные субъединицы ведут к полной инактивации всех стадий транскрипции. α субъединица необходима для ассоциации белков в холофермент. Когда фаг Т4 инфицирует E.coli, альфа субъединица подвергается АДФ-рибозилированию по остатку аргинина. Эта модификация связанна с сильным падением сродства фермента к промоторам, которые до этого узнавались холоферментом, что доказывает участие этой субъединицы в узнавании промотора. Так же альфа субъединица играет важную роль в процессах взаимодействия РНК-полимеразы с различными регуляторными белками.

8
Строение РНК-полимеразы:
δ субъединица отвечает за специфическое узнавание промотора и просоединение к нему других белков РНК-полимеразы. Существуют так же очень маленькие РНК-полимеразы, представляющие собой единственную полипептидную цепь, кодируемую единственным фаговым геном. Это говорит о том, что необходимый для транскрипции генов фермент может быть гораздо меньшим, чем многобелковый комплекс клетки-хозяина. Таким образом можно говорить о неком минимальном комплексе РНК-полимеразы. Но такой белок узнает только некоторые промоторы фаговой ДНК и не имеет таких широких возможностей для смены промоторов, как хозяйская РНК-полимераза.

9
Таким образом холофермент можно разделить на два компонента кор-фермент и сигма-фактор.
Только холофермент может инициировать транскрипцию. Сигма-фактор обеспечивает прочное связывание РНК-полимеразы с промотором ДНК. Сигма-фактор освобождается, когда будут синтезированы первые 8-9 оснований в цепи РНК, а кор-фермент продолжит процесс элонгации.

10
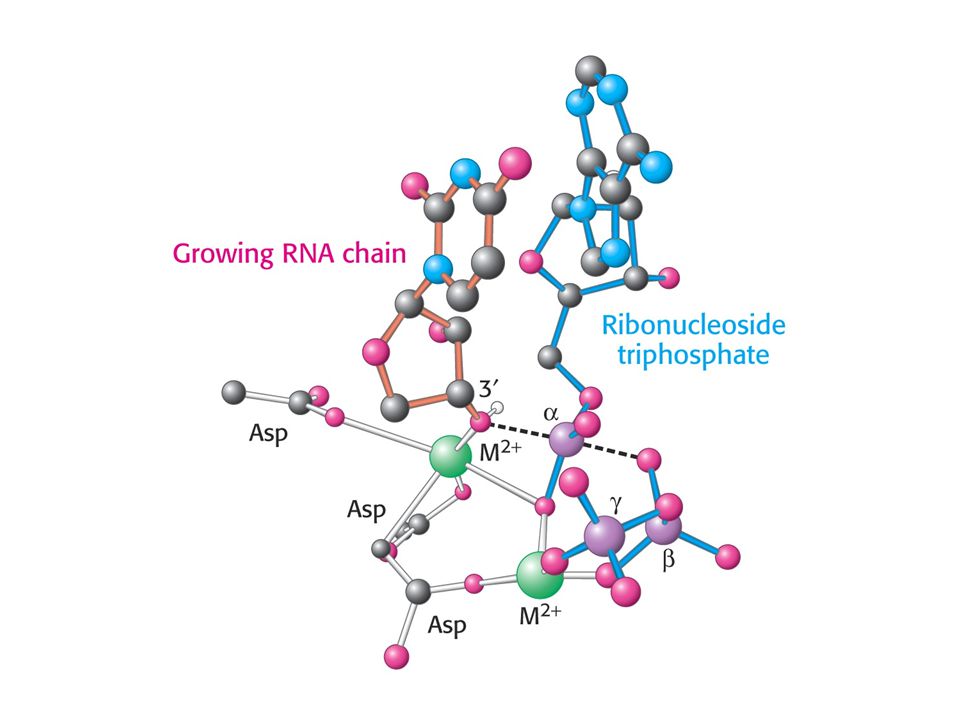
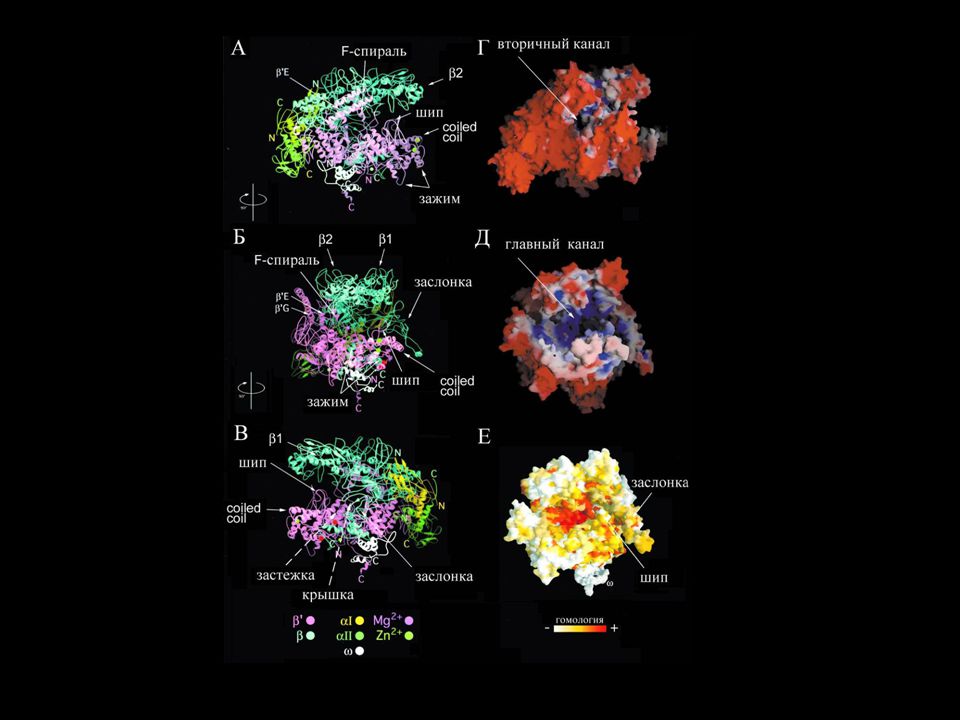
Структура кор-фермента РНК-полимеразы
Вторичный канал Главный канал Mg

11
Структурная модель элонгационного комплекса
шип зажим 5’-РНК 3’-РНК заслонка вторичный канал (НТФ)
")
12
Функцональные активности РНКП
РНКП способна катализировать пять различных химических реакций, а именно: Синтез цепи РНК на матрице ДНК. Экзопирофосфоролиз Реакция экзонуклеазного расщепления новосинтезированного РНК-транскрипта Эндопирофосфоролиз Реакция эндонуклеазного расщепления новосинтезированного РНК-транскрипта

20
Промоторы: Транскрипция инициируется при образовании стабильного комплекса между холоферментом и специфической последовательностью, называемой промотором и располагающейся в начале всех транскрипционных единиц. Изучение нуклеотидной последовательности более чем 50 разных промоторных сайтов прокариот и мутационный анализ выявили только два консервативных участка, по-видимому играющих ключевую роль в узнавании и функционировании промотора.

21
Прибнов-бокс (-10 элемент):
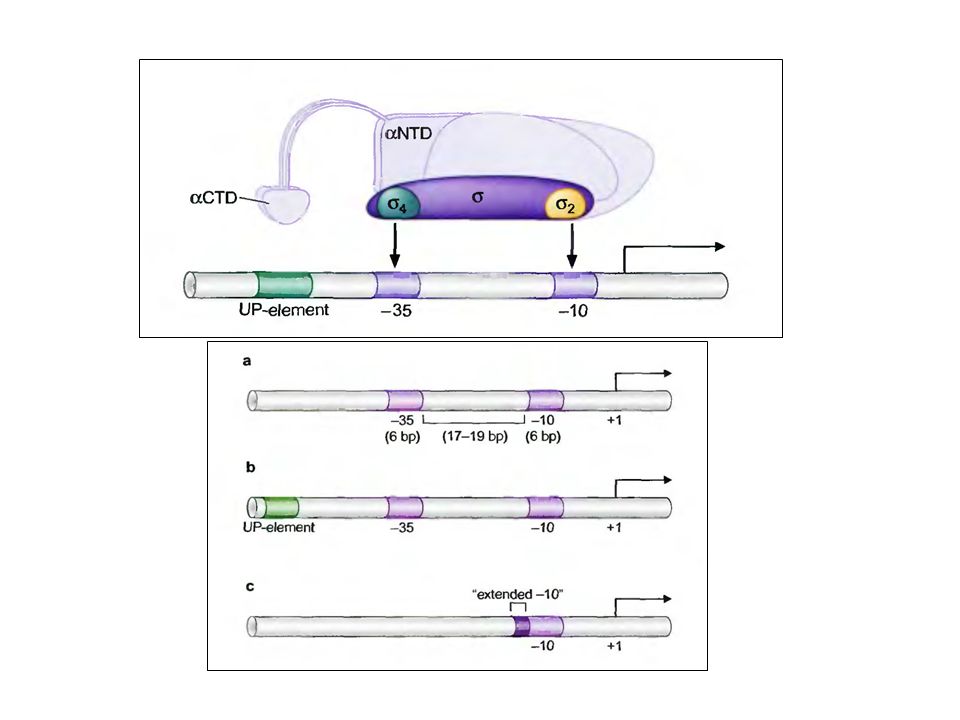
Одна из этих последовательностей состоит из шести или семи пар оснований и расположена на расстоянии примерно 10 оснований до того нуклеотида, с которого начинается транскрипция (+1); этот сигнал обычно обозначают как — 10-последовательность, или Прибнов-бокс-в честь ее открывателя. Сравнительный анализ — 10-последовательностей примерно 50 промоторов прокариот показал, что все они немного отличаются от консенсус-последовательности ТАТААТ.
; этот сигнал обычно обозначают как — 10-последовательность, или Прибнов-бокс-в честь ее открывателя. Сравнительный анализ — 10-последовательностей примерно 50 промоторов прокариот показал, что все они немного отличаются от консенсус-последовательности ТАТААТ.")
22
-35 элемент: Вторая последовательность, длина которой обычно равна девяти нуклеотидам, расположена на расстоянии 35 оснований до сайта инициации ( — 35-последовательность) и также встречается в большинстве промоторов прокариот. Нуклеотидная последовательность сегмента между —35- и —10-участками не является критической, важно лишь расстояние между этими участками —35-последовательность участвует в связывании РНК-полимеразы, которое предшествует перемещению фермента в Прибнов-бокс.
 и также встречается в большинстве промоторов прокариот. Нуклеотидная последовательность сегмента между —35- и —10-участками не является критической, важно лишь расстояние между этими участками —35-последовательность участвует в связывании РНК-полимеразы, которое предшествует перемещению фермента в Прибнов-бокс.")
25
3 2 1 4 2 α 2 Промоторные элементы бактерий Холо- фермент
Кор-фермент 2 2 1 4 2 TATAAT TTGACA TATAAT UP -35 -10 Некоторые сильные промоторы (например, промоторы рибосомальных РНК) содержат дополнительный UP-элемент (от upstream) левее -35 области… …который узнается -субъединицей РНКП
 содержат дополнительный UP-элемент (от upstream) левее -35 области… …который узнается -субъединицей РНКП.")
27
Принцип регуляции 1 Различия в эффективности транскрипции индивидуальных генов отчасти зависят от структуры их промоторов. Как прочность взаимодействия РНК-полимеразы с промоторной последовательностью, так и эффективность образования «открытого» про-моторного комплекса определяются конкретными нуклеотидными последовательностями -35 и -10-участков соответственно. Мутации в этих участках приводят к значительным изменениям способности многих промоторов обеспечивать инициацию транскрипции.

28
Принцип регуляции 2 Существует разделение функций между кор-ферментом, занимающимся элонгацией цепи РНК и сигма-фактором, сайт-специфично узнающим последовательности промоторов ДНК. В клетке возможно наличие нескольких различных сигма субъединиц, узнающих различные типы промоторов. Смена сигма-факторов часто происходит при кардинальных перестройках экспрессии генов, например в ответ на изменение образа жизни бактерии при переходе к споруляции или сильном изменении условий окружающей среды.

29
Альтернативные δ-факторы:
Основной фактор, ответственный за транскрипцию большинства клеточных генов в нормальных условиях называется δ70. Альтернативные сигма-факторы δЕ, δ32,δ54 активируются в ответ на изменения окружающей среды, а сигма δ28 служит для экспрессии генов белков бактериальных жгутиков (флагелл), в течении нормальной жизнедеятельности клетки, но в ответ на изменения среды. Все сигма-факторы, кроме δ54 принадлежат к одному белковому семейству с общим строением и происхождением.
, в течении нормальной жизнедеятельности клетки, но в ответ на изменения среды. Все сигма-факторы, кроме δ54 принадлежат к одному белковому семейству с общим строением и происхождением.")
30
Регуляция транскрипции у прокариот
Большинство бактерий содержат несколько -субъединиц, которые отвечают за узнавание разных типов промоторов и транскрипцию различных групп генов, объединенных какой-то общей функцией. Escherichia coli: 70 – гены “домашнего хозяйства” 32 – тепловой шок 38 – стрессовые условия плавление промотора без затрат АТФ зависит от суперспирализации 54 – условия азотного голодания плавление промотора при участии белка-активатора с затратами АТФ

31
Тепловой шок: Все прокариоты и эукариоты имеют особую систему белков, которая начинает интенсивно экспрессироваться в ответ на тепловой стресс. При возрастании температуры синтезируется особый набор белков, а синтез основных белков за счет узнавания промотора сигма 70 снижается. Белки этого набора являются продуктами экспрессии генов белков теплового шока. Эти белки защищают клетку от последствий теплового шока. Например E.coli экспрессирует 17 подобных белков за счет переключения сигма субъединиц, узнающих разные промоторы. Ген rpoH абсолютно необходим для подобного переключения, его продуктом является сигма32, функционирующая как альтернативный сигма-фактор. Интенсивность экспрессии белков теплового шока растет пропорционально росту температуры. Главным стимулом к переключению сигма-факторов служит накопление несвернутых и различно поврежденных белков внутри клетки.

32
Тепловой шок: Другая группа генов теплового шока контролируется сигма Е. Эти гены экспрессируются в ответ на более экстремальные изменения температур, чем при синтезе сигма32. Здесь индуктором к синтезу также служит наличие различно денатурированных белков в цитоплазме и мембранах.

33
Азотное голодание: Другой сигма-фактор используется в условиях азотного голодания. Клетки E.coli содержат другую минорную сигма субъединицу – сигма 54, которая активируется когда в клетке возникает недостаток ионов аммония. Таким образом становиться возможной экспрессия генов, отвечающих за использование альтернативных источников азота. Функциональные двойники этой сигма субъединицы были найдены во многих бактериальных клетках, что говорит о неком эволюционно общим механизме борьбы с азотным голоданием.

34
Вывод: Таким образом при замене одной сигма субъединицы на другую происходит изменение экспрессии целого набора генов, промотор которых способен специфически связываться только с определенным сигма-фактором.

36
Стадии инициации: При описании процесса инициации транскрипции выделяют ряд различных стадий, в зависимости от состояния РНК-полимеразы и ДНК. Реакция взаимодействия холофермента с промотором начинается с формирования закрытого комплекса. Образование этого комплекса будет обратимым. Закрытый комплекс переходит в открытый при плавлении короткой последовательности ДНК, с которой связана РНК-полимераза. Для сильных промоторов эта стадия полностью необратима, а реакция перехода проходит очень быстро.

37
Стадии инициации: Следующей стадией считается включение первых двух нуклеотидов с формированием фосфодиэфирной связи между ними. Это обеспечивает формирование прочного комплекса, включающего в себя РНК, ДНК и фермент. Дальнейшие нуклеотиды могут добавляться без продвижения фермента вдоль матрицы, формируя короткие цепи длиной в 9 нуклеотидов. Но фермент может здесь обрывать синтез цепи, высвобождая девяти нуклеотидные цепи. Этот процесс называется абортивной инициацией, после которой фермент начинает синтез цепи РНК заново. Когда инициация прошла успешно, фермент теряет сигма субъединицу и переходит в прочный элонгационный комплекс. Это произойдет, когда фермент синтезирует цепь длиннее 10 нуклеотидов и тем самым начнет продвигаться по матрице ДНК.

38
Элонгация: В течение периода элонгации фермент продвигается вдоль ДНК и удлиняет растущую цепь ДНК. Фермент продвигается раскручивая дуплекс ДНК и таким образом освобождая матрицу для построения на ней растущей РНК. Нуклеотиды ковалентно добавляются к 3’ концу растущей РНК, формируя ДНК-РНК гибридный дуплекс. Затем цепи расплетенного дуплекса ДНК снова ассоциируют, восстанавливая исходную структуру. Одноцепочечная РНК выходит из активного центра полимеразы.

39
Элонгационный комплекс
Активный центр РНКП способен расщеплять РНК: 1. Исправление ошибок 2. Реактивация «смещенных» комплексов 3’-РНК NTP Синтез РНК GreA, GreB Смещение назад 3’

40
Арест элонгации: В некоторых случаях могут возникать паузы в процессе элонгации, например в результате нехватки предшественников нуклеотидов при проведении реакции в бесклеточной системе. Подобные паузы называют арестом элонгации. Когда нехватка предшественников устраняется, то фермент преодолевает данную остановку путем внесения особого 3’-разрыва в синтезированную нить РНК, это позволяет создать новый 3’-конец цепи для ее далнейшей элонгации. За подобное расщепление ответственны особые белки GreA и GreB, освобождающие РНК-полимеразу от элонгационного ареста.

41
Стадии транскрипции: Терминатор – это последовательность ДНК, означающая окончание транскрипции, в результате чего РНК-полимераза прекращает транскрипцию и диссоциирует с матрицы. Основным событием терминации является узнавание особой точки, в которой синтез ДНК должен завершиться. Когда последнее основание будет присоединено к цепи РНК, транскрипционный глазок разрушается: гибридный дуплекс ДНК-РНК плавится, восстанавливается двухцепочечная структура района ДНК, фермент диссоциирует с матрицы.

42
Терминация: Однажды РНК-полимераза начинает транскрипцию, продвигаясь вдоль дуплекса ДНК до тех пор, пока не встретит особую последовательность терминатора. В этой точке фермент заканчивает добавлять нуклеотиды к растущей цепи РНК и диссоциирует с матрицы, высвобождая первичный транскрипт. Терминация обязательно должна включать стадию разрыва водородных связей в дуплексе ДНК-РНК и восстановления исходной структуры ДНК

43
Два типа терминаторов:
Кор-фермент может терминировать в опытах в некоторых сайтах, при отсутствии специфических дополнительных белков. Такие сайты будут называться внутренними терминаторами. Rhо-зависимый терминатор определяется необходимостью добавления ро-фактора в эксперименте, и мутации по этому белку доказывают его необходимость для терминации в живой клетке.

44
Механизмы терминации -независимая терминация -зависимая терминация
Синтезиуемая РНК способна образовывать различные вторичные структуры, что может иметь важное регуляторное значение

45
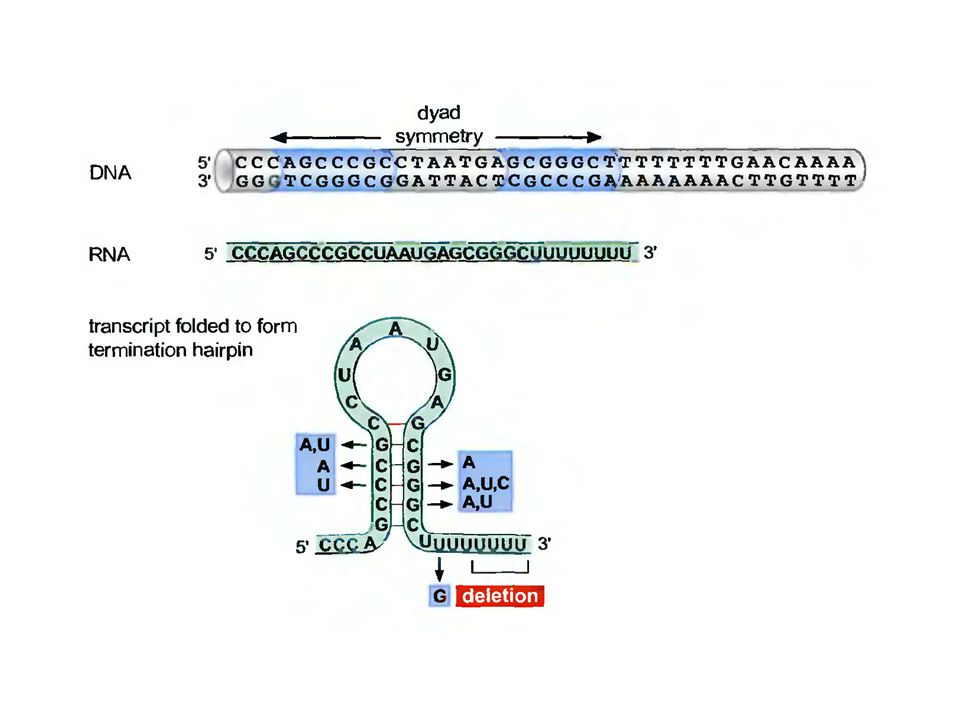
Внутренний терминатор:
Внутренний терминатор имеет два структурных мотива абсолютно необходимых для успешной терминации: участок вторичной структуры – шпилька последовательность остатков уридина на самом конце этой шпильки. Шпилька часто содержит GC- богатый регион недалеко от основания стебля. Мутации ингибирующие терминацию на данном типе последовательностей локализуются в районе ствола шпильки РНК.

47
Механизм работы: Возможно, что все шпильки, которые образуются при траскрипции гена, создают затруднения для дальнейшего продвижения РНК-полимеразы, благодаря чему возникают короткие паузы в ее работе. Уридин-богатый гибрид РНК-ДНК содержит очень непрочные водородные связи, из-за чего вся структура становится непрочной и возможна легкая диссоциация подобного дуплекса. При паузах на шпилечной структуре легко происходит плавление подобного дуплекса и освобождение первичного транскрипта.

48
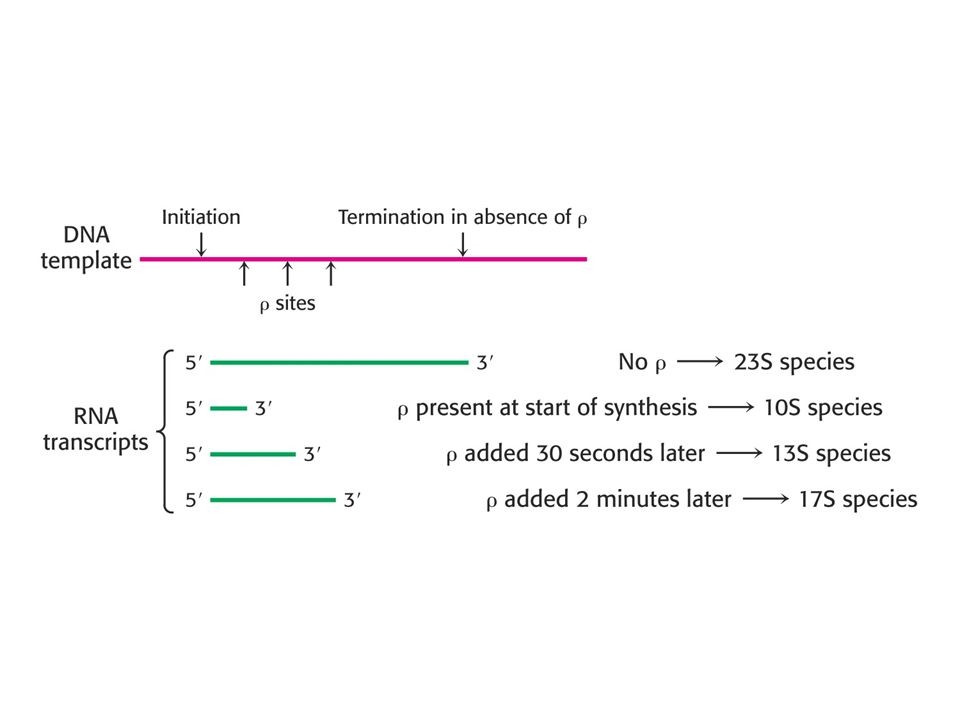
Ро-зависимая терминация:
Ро-фактор так же необходим для успешной терминации транскрипции различных клеточных генов. Он функционирует исключительно на стадиях терминации. Ро-фактор представляет собой гексамер весом 275 кДа, состоящий из одинаковых субъединиц. Ро-зависимый терминатор представляет собой последовательность нуклеотидов богатую цитозином и бедную гуанином. E.coli содержит относительно немного ро-зависимых терминаторов, большинство из них характерно для различных фаговых геномов.

50
Механизм работы: Rho-фактор обладает АТФазной активностью, которая требует наличия полирибонуклеотида. Это доказывает, что rho-фактор связывается с РНК и может по ней передвигаться. Существует так называемая модель «горячего преследования» rho-фактором РНК-полимеразы, в которой ро-фактор связывается с особыми сайтами растущей нити РНК или просто с какими-то ее выступающими частями, и следует по ней за ферментом. Возможно далее rho-фактор передвигается по РНК быстрее, чем полимераза по ДНК и если фермент делает паузу, когда встречается с терминатором, то rho-белок просто вытесняет его с матрицы, захватывая последовательность РНК-терминатора. Так же в отсутствии РНК-полимеразы, rho-фактор проявляет геликазную активность расплетая дуплекс РНК-ДНК.

52
Регуляция работы генов
Транскрипция Регуляция работы генов

53
Азотное голодание Индикатором количества биологически связанного азота в клетке является соотношение концентраций глутамина и кетоглутарата. При недостатке азота в клетке начинает накапливаться кетоглутарат, взаимодействующий с регуляторным белком UTase/UR, способным в присутствии кетоглутарата уридилировать PII белок. В присутствии глутамата, наоборот, активируется способность UTase/UR снимать остаток уридиловой кислоты с PII. Неуридилированная форма белка PII способна связывать регуляторный белок NtrB и активировать его фосфатазную активность. Эта активность направлена на ДНК-связывающий белок NtrC. При избытке азота в клетке информация, в виде каскада химических модификаций приводит к немодифицированной (дефосфорилированной) форме NtrC. При недостатке азота, наоборот, свободный (от PII) NtrB переносит фосфат с ATP на NtrC. Фосфорилированный NtrC взаимодействует с комплексом РНК-полимеразы с s54, который находится на промоторах генов участвующих в метаболизме азота.
 форме NtrC. При недостатке азота, наоборот, свободный (от PII) NtrB переносит фосфат с ATP на NtrC. Фосфорилированный NtrC взаимодействует с комплексом РНК-полимеразы с s54, который находится на промоторах генов участвующих в метаболизме азота.")
54
При недостатке азота происходит активация транскрипции генов, необходимых для восстановления азотного баланса. Данный пример показывает два механизма, широко использующихся для регуляции белковых систем: Во-первых, использование ковалентной модификации, например фосфорилирования, для изменения активности белков. Во-вторых, использование многоступенчатых “каскадов” регуляторных белков. Такие каскады необходимы для усиления сигнала. Т.к. один активированный первичный регуляторный фермент (в данном случае UTase/UR) может модифицировать много PII белков, а в свою очередь каждый модифицированный PII в комплексе с NtrB активирует много NtrC. Кроме того, ступенчатые каскады позволяют принимать “сигналы” от многих других регуляторных систем.
 может модифицировать много PII белков, а в свою очередь каждый модифицированный PII в комплексе с NtrB активирует много NtrC. Кроме того, ступенчатые каскады позволяют принимать сигналы от многих других регуляторных систем.")
56
Регуляция экспрессии лактозного оперона
Негативная регуляция

57
Регуляция транскрипции у прокариот
Бактериальные гены организованы в опероны типичный участок генома бактерий Оперон - группа генов, транскрибируемых в составе одной РНК; регулируются совместно и обычно обладают общей функцией. Регулон - группа совместно регулируемых оперонов, обладающих какой-либо общей функцией

58
(например, лактоза, галактоза)
Регуляция транскрипции у прокариот Активация Репрессия оператор оператор оператор – участок связывания регуляторного белка связывание активаторов и репрессоров с операторами контролируется какими-либо клеточными метаболитами-индукторами (например, лактоза, галактоза) Активаторы обычно привлекают РНКП к промотору Репрессоры обычно мешают связыванию РНКП с промотором
 Активаторы обычно привлекают РНКП к промотору. Репрессоры обычно мешают связыванию РНКП с промотором.")
59
Общие сведения: Бактерии Е. соli могут использовать в качестве единственного источника углерода и энергии лактозу, поскольку они способны образовывать в большом количестве галактозидазу - фермент, расщепляющий лактозу на глюкозу и галактозу. Однако при росте на других источниках углерода в клетках Е.соli образуется очень мало-галактозидазы. Ген, ответственный за синтез галактозидазы (lасZ), называется индуцибельным, поскольку кодируемый им фермент синтезируется только тогда, когда в клетке присутствуют сахара, имеющие галактозильные остатки.
, называется индуцибельным, поскольку кодируемый им фермент синтезируется только тогда, когда в клетке присутствуют сахара, имеющие галактозильные остатки.")
60
Структура lac-оперона:
Помимо галактозидазы может быть индуцировано образование еще двух белков: галактозидпермеазы (кодируемой геном lасY), необходимой для проникновения бета-галактозидов в клетку, и бета-галактозидтрансацетилазы (lасА), фермента с окончательно невыясненной пока функцией. В этих трех генах-lасZ, lасY и lасА - содержится вся информация о белках, кодируемых lас-опероном. Они транскрибируются в единую полицистронную РНК, при трансляции которой образуются почти одинаковые количества соответствующих белков. Поэтому можно сказать, что три гена lас-оперона экспрессируются согласованно. Со структурными генами lас-оперона связаны несколько типов регуляторных элементов, ответственных за индуцибельность и координированную регуляцию этих генов.
, необходимой для проникновения бета-галактозидов в клетку, и бета-галактозидтрансацетилазы (lасА), фермента с окончательно невыясненной пока функцией. В этих трех генах-lасZ, lасY и lасА - содержится вся информация о белках, кодируемых lас-опероном. Они транскрибируются в единую полицистронную РНК, при трансляции которой образуются почти одинаковые количества соответствующих белков. Поэтому можно сказать, что три гена lас-оперона экспрессируются согласованно. Со структурными генами lас-оперона связаны несколько типов регуляторных элементов, ответственных за индуцибельность и координированную регуляцию этих генов.")
61
Основная идея: Поскольку промоторная и операторная последовательности перекрываются, связывание репрессора с оператором мешает связыванию РНК-полимеразы с промотором, что приводит к блокированию транскрипции структурных генов. Транскрипцию оперона можно индуцировать, если блокировать связывание репрессора с оператором. Такое блокирование происходит при связывании одного из галактозидов с той или иной субъединицей репрессора, что уменьшает сродство последнего к оператору. После отсоединения репрессора от промотра полимераза может связаться с промотором и инициировать транскрипцию оперона.

62
Регуляция экспрессии лактозного оперона
Позитивная регуляция

63
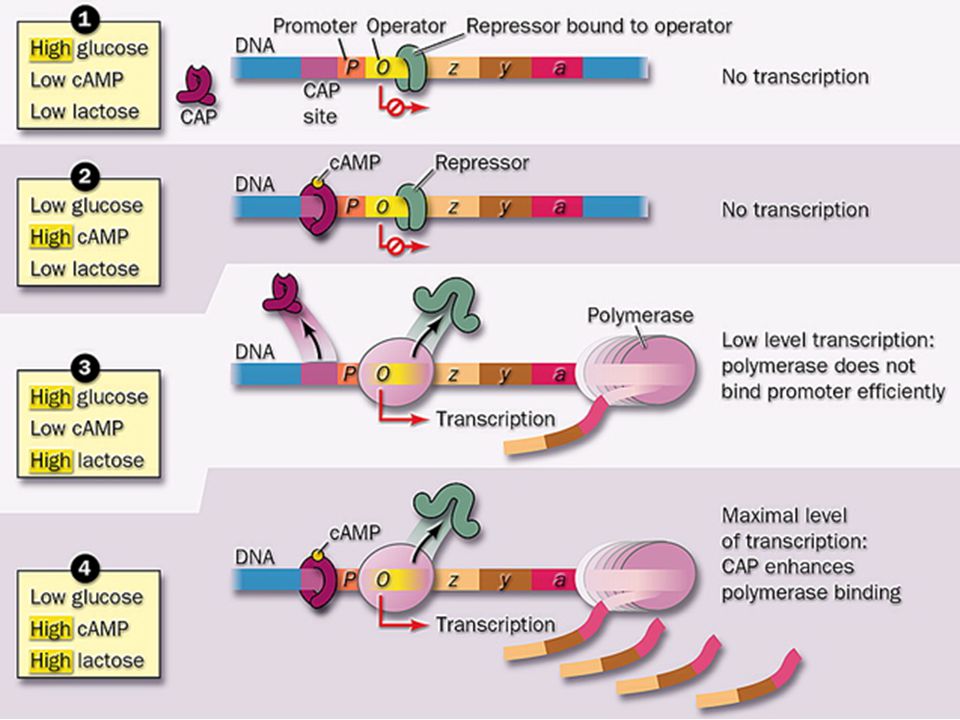
Общие сведения: Для экспрессии lас-оперона, как и других индуцибельных оперонов, которые осуществляют контроль синтеза ферментов, участвующих в метаболизме cахаров, необходимо не только снять репрессию оперона, но и получить некий сигнал. Таким сигналом служит комплекс циклического АМР (сАМР) с белком-активатором катаболизма (САР, от англ. Саtabolite activator protein), который связывается со специфической последовательностью, находящейся в самом начале lас-промотора. сАМР, принимающий участие во многих клеточных процессах, образуется из АТР в ответ на самые разные вне- и внутриклеточные события.
 с белком-активатором катаболизма (САР, от англ. Саtabolite activator protein), который связывается со специфической последовательностью, находящейся в самом начале lас-промотора. сАМР, принимающий участие во многих клеточных процессах, образуется из АТР в ответ на самые разные вне- и внутриклеточные события.")
64
Механизм влияния: САР представляет собой димер из идентичных полипептидных цепей с мол. массой 22 кДа. Связывание комплекса САР-сАМР со специфической последовательностью в начале промотора приводит к усилению транскрипции lас-оперона почти в 50 раз. Сам по себе САР не способен к такому связыванию и стимуляции транскрипции. Усиление транскрипции с помощью комплекса САР-сАМР можно объяснить тем, что, связываясь с ДНК в непосредственной близости от сайта присоединения РНК-полимеразы, он усиливает сродство этого фермента к промотору. Альтернативная гипотеза заключается в том, что связывание САР-сАМР с САР-сайтом предотвращает присоединение РНК-полимеразы к расположенному поблизости слабому промотору и увеличивает тем самым вероятность того, что полимераза свяжется с «правильным» промоторным сайтом.

66
Регуляция транскрипции у прокариот
Узнавание пар нуклеотидов аминокислотами Узнавание специфической последовательности ДНК белками helix-turn-helix

67
Регуляция транскрипции у прокариот
СAP – catabolite activator protein cAMP (синтезируется в отсутствие глюкозы) Взаимодействие с -субъединицей РНКП Активация катаболитных оперонов TGTGA ACACT TCACA AGTGT
 Взаимодействие с -субъединицей РНКП. Активация катаболитных оперонов. TGTGA. ACACT. TCACA. AGTGT.")
69
Регуляция транскрипции у прокариот
Связывание белка-активатора Удаление белка-репрессора СAP R R Активация специфического оперона индуктор R

70
Реитеративная транскрипция
Известно, что при инициации транскрипции первые несколько полимеризованных нуклеотидов плохо связаны в транскрипционном комплексе. Эта особенность используется при регуляции транскрипции пиримидинового оперона. Если концентрация UTP в клетке высока, нет нужды транскрибировать этот оперон. В начале оперона расположен короткий участок, кодирующий олигоуридиловую кислоту. При высокой концентрации UTP олигоU фрагмент начинает проскальзывать по ДНК, так, что синтезируется длинные олигоуридиловые фрагменты, что приводит к терминации транскрипции

71
Регуляция терминации Кроме стадии инициации, транскрипция может регулироваться также с помощью преждевременной терминации, т.е. в зависимости от нужд клетки должен осуществляться выбор между элонгацией и терминацией. Наиболее простой способ остановить транскрипцию – связать белок-регулятор на пути РНК-полимеразы. Так происходит в случае оперона, ответственного за синтез пуриновых нуклеотидов. Два более сложных механизма регулируемой терминации это антитерминация и аттенюация.

72
Аттенюация Аттенюация – это регулируемая терминация зависящая от относительной стабилизации альтернативных вторичных структур транскрибируемой мРНК. Как известно, r-независимый терминатор представляет собой РНК шпильку и последующий олигоуридиловый участок. Если будет стабилизирована альтернативная вторичная структура, где олигоуридиловый фрагмент удален от ближайшей шпильки, терминация не будет происходить. Есть несколько факторов, стабилизирующих одну из альтернативных вторичных структур РНК во время транскрипции.

73
Триптофановый оперон Транскрипция триптофанового оперона E. coli должна происходить только когда концентрация триптофана в клетке мала. В начале trp-оперона закодирован короткий пептид, богатый остатками триптофана. Поскольку трансляция мРНК у кишечной палочки происходит котранскрипционно, лидерный участок мРНК начинает транслироваться сразу после синтеза. Для лидерного участка мРНК существуют две альтернативные вторичные структуры, одна способствующая преждевременной терминации, другая нет. Если рибосома быстро транслирует лидерный участок мРНК, что характерно для высоких концентраций триптофана, она расплавляет элемент вторичной структуры, препятствующий терминации и, таким образом, стабилизирует вторичную структуру терминатора.

74
Регуляция транскрипции у прокариот: аттенюация
гены биосинтеза Trp нехватка Trp достаточно Trp

75
Рибопереключатели

76
Регуляция транскрипции у прокариот: рибопереключатели (riboswitches)
Инициация транскрипции Инициация трансляции В присутствии метаболита: образуется структура терминатора транскрипции маскируется участок связывания с рибосомой

77
А Б

78
Рибопереключатели А Б

80
Регуляция транскрипции у прокариот: рибозимы
Рибозимы – молекулы РНК, обладающие каталитической активностью Консервативный элемент в 5’-концевой области гена glmS B. subtilis: В присутствии глутамина происходит активация рибозима и расщепление РНК

81
Антитерминация Лучше всего процесс антитерминации изучен для бактериофага l. У данного фага имеется два механизма антитерминации. Оба механизма связаны с регуляторными белками-антитерминаторами. Белок N фага l связывается с определенной структурой на транскрибируемой РНК – nut (от N utilization). На данной структуре при участии N собирается РНК-белковый комплекс, включающий белки S10, NusA, NusB и NusG, взаимодействующий с РНК-полимеразой. Этот комплекс препятствует терминации данного РНК-полимеразного комплекса на r-зависимых и некоторых r-независимых терминаторах.
. На данной структуре при участии N собирается РНК-белковый комплекс, включающий белки S10, NusA, NusB и NusG, взаимодействующий с РНК-полимеразой. Этот комплекс препятствует терминации данного РНК-полимеразного комплекса на r-зависимых и некоторых r-независимых терминаторах.")
82
Антитерминация По-другому устроен механизм антитерминации при помощи белка Q фага l. Это ДНК-связывающий белок, место связывания которого располагается с 5’-стороны от промотора оперона, регулируемого этим белком. Сразу за точкой начала транскрипции расположен сильный r-независимый терминатор, активность которого подавляется белком Q.

83
Для регуляции транскрипции используются как стадии инициации, так и элонгации/терминации.
Замена основного фактора инициации s используется в случае, когда необходимо существенно изменить спектр белков, синтезируемых в клетке. Для более тонкой регуляции используются регуляторные ДНК связывающие белки активаторы и ингибиторы. ДНК-связывающая способность этих белков определяется связыванием лигандов или ковалентной модификацией. На стадии элонгации и терминации регуляция может осуществляться с помощью аттенюации, когда мРНК может иметь альтернативные вторичные структуры, одна из которых способствует терминации. Относительная стабилизация какой-либо из вторичных структур определяется либо скоростью транскрипции, либо скоростью трансляции, либо связыванием белка. Антитерминация, или подавление активности некоторых терминаторов может происходить с помощью регуляторных антитерминирующих белков.

84
Регуляция транскрипции фага лямбда
Наиболее хорошо изученный объект исследования регуляции транскрипции – бактериофаг l. Для него существует два альтернативных пути развития. Если ресурсов клетки много, фаг избирает литический путь развития, т.е. старается синтезировать как можно больше дочерних фаговых частиц и в конце концов лизирует клетку. Если ресурсов клетки, зараженной l мало, то бактериофаг предпочитает интегрироваться в геном клетки и переждать “трудные времена” в лизогенном состоянии. Когда клетке грозит опасность, такой интегрированный профаг стремиться “бежать с тонущего корабля” и активирует литическую программу развития.

87
Первые активные промоторы при попадании l в клетку - PL и PR.
Транскрипты с этих промоторов ограничиваются терминаторами tL1 и tR1, т.е синтезируются белки N и cro. Белок N является антитерминаторным белком и позволяет РНК полимеразе транскрибировать гены CII и CIII. Тетрамерный СII белок является регуляторным и активирует ряд промоторов фага l, помеченных символом “/CII”. Белок CII нестабилен и легко подвергается протеолизу клеточными протеазами HflA и HflB. Стабильность CII определяет то, литический или лизогенный путь развития выберет фаг l. Если клетке не хватает энергетических ресурсов, что на молекулярном уровне приводит к накоплению cAMP и активации белка CAP, репрессируется активность HflA и, соответственно, стабилизируется активатор лизогенного состояния – CII.

88
Лизогения Стабилизация CII приводит к активации промоторов PI/CII, PRE/CII и PaQ/CII. Таким образом, синтезируются антисмысловые РНК к мРНК регуляторного белка cro и белков O и P, участвующих в репликации l. Синтезируются интеграза Int и регуляторный белок CI. После интеграции в геном клетки, синтезируется только белок CI, активирующий свой собственный промотор PRM и ингибирующий промоторы PL и PR.

89
Побег из генома При индукции в клетке SOS-системы, т.е. когда клетке угрожает опасность, клеточный белок RecA индуцирует разрезание репрессора SOS-системы lexA. Фаг лямбда использует этот механизм, сделав CI также подверженным RecA-индуцированному разрезанию. При деструкции CI включаются PR и PL промоторы. Появляющийся белок cro инактивирует PRM и способствует активации PR и PL, делая процесс литического развития необратимым.

90
Лизис При продвижении фага по литическому пути подавлена активность промоторов, активируемых CI и CII. Синтезируются белки репликазного комплекса O и P, начинает синтезироваться антитерминаторный белок Q. На последнем этапе литического каскада Q выключает терминатор tt. Таким образом, с промотора PR начинают синтезироваться белки оболочки и лизиса.

91
Регуляция транскрипции фага лямбда иллюстрирует то, как комбинацией действия различных белков-регуляторов можно достигать точно упорядоченной во времени экпрессии генов. Фаг лямбда также может служить примером того, как различные генетические программы включаются и выключаются в зависимости от необходимости.

92
Регуляция транскрипции фага T4
Вместе с ДНК фага T4 в клетку проникает белок Alt, ADP-рибозилирующий a-субъединицу РНК-полимеразы. Эта модификация препятствует взаимодействию РНК-полимеразы с наиболее активными промоторами клетки, содержащими специфическую активаторную последовательность. В тоже время промоторы фага T4 лучше узнаются модифицированной a-субъединицей. Ранние гены фага T4 транскрибируются с использованием обычного клеточного s-фактора. Среди этих ранних генов несколько отвечают за активацию средних генов и, одновременно, выключение уже всех промоторов клетки-хозяина.

93
Белок Mod завершает модификацию a-субъединиц РНК-полимеразы
Белок Mod завершает модификацию a-субъединиц РНК-полимеразы. Белок Alc связывается с цитозинсодержащими последовательностями клеточной ДНК и вызывает преждевременную терминацию транскрипции. С ДНК фага T4 этот белок не связывается, поскольку вместо цитозина эта ДНК содержит 5-оксиметилцитозин. Вирусный белок AsiA связывается с s-фактором и переключает его специфичность с клеточной промоторной –35 последовательности на активаторный белок бактериофага T4 – MotA. Белок MotA узнает mot-последовательность, функциональный аналог –35 участка для промоторов средних генов.

94
При активации поздних генов бактериофаг полностью отказывается от клеточного s-фактора и использует вместо него белок gp55, узнающий промоторы поздних генов. Адаптором между gp55 и РНК-полимеразой служит gp33. Кроме того, энхансером транскрипции поздних генов T4 служат компоненты репликативного комплекса этого фага. Фактор процессивности репликации – тример gp45 активирует транскрипцию поздних генов при помощи комплекса gp44/gp62.

Similar presentations
















 Ian Sommervillle Часть 4. Реализация ПО: Проектирование с повторным использованием компонентов.>")


 (2) (3) (4) (5) (6) (7) Математическое моделирование процессов отбора2.>")





 Томск, 2011 НИИ фармакологии СО РАМН ИСЭ СО РАН.>")


 917-23-89 119019 Москва, а/я 212 Рабочая группа по реформе МВД Москва, 2010 Новикова Асмик, Фонд «Общественный вердикт»>")

