Download presentation
Presentation is loading. Please wait.

1
Final Exam Mon 8 am

2
Lecture Connections 22 | Nitrogen Assimilation, Biosynthetic Use, and Excretion © 2009 W. H. Freeman and Company

3
CHAPTER 22 Nitrogen Assimilation, Biosynthetic Use, and Excretion –Nitrogen fixation –Incorporation of ammonia into biomolecules –Biosynthesis of amino acids –Biosynthesis of heme –Biosynthesis of nucleotides –Catabolism of purines Key topics:

4
Importance of Nitrogen in Biochemistry Nitrogen, carbon, hydrogen and oxygen are the main elemental constituents of living organisms Peptide backbone in proteins Functional side chains (His, Lys, Arg, Trp, Asn, Gln) in proteins Nucleobases in DNA and RNA Found in several cofactors (NAD, FAD, Biotin … ) Found in many small hormones (epinephrine) Found in many neurotransmitters (serotonin) Found in many pigments (chlorophyll) Found in many defense chemicals (amanitin)
 in proteins Nucleobases in DNA and RNA Found in several cofactors (NAD, FAD, Biotin … ) Found in many small hormones (epinephrine) Found in many neurotransmitters (serotonin) Found in many pigments (chlorophyll) Found in many defense chemicals (amanitin)")
5
Biochemistry of Molecular Nitrogen Our atmosphere is rich in nitrogen but its chemical inertness prevents use of N 2 by most organisms Atmospheric nitrogen is converted to nitrogen compounds by a few non-biological processes –NO from N 2 and O 2 during lighting –NH 3 from N 2 and H 2 in the industrial Haber process Atmospheric nitrogen is fixed directly by certain bacteria and archae –Most are free-living single-celled prokaryotes (archaea) –Some live in symbiosis with plants (e.g. proteobacteria with legumes) –Few live in symbiosis with animals (e.g. spirochaete with termites)
 –Few live in symbiosis with animals (e.g. spirochaete with termites).")
6
Nitrogen-fixing nodules. (a) Root nodules of bird's-foot trefoil, a legume. (b) Artificially colorized electron micrograph of a thin section through a pea root nodule. Symbiotic nitrogen-fixing bacteria, or bacteroids, live inside the nodule cell, surrounded by the peribacteroid membrane. Bacteroids produce the nitrogenase complex that converts atmospheric nitrogen (N 2 ) to ammonium (NH 4 + ); without the bacteroids, the plant is unable to utilize N 2. The infected root cell provides some factors essential for nitrogen fixation, including leghemoglobin; this heme protein has a very high binding affinity for oxygen, which strongly inhibits nitrogenase. nucleus
 Artificially colorized electron micrograph of a thin section through a pea root nodule. Symbiotic nitrogen-fixing bacteria, or bacteroids, live inside the nodule cell, surrounded by the peribacteroid membrane. Bacteroids produce the nitrogenase complex that converts atmospheric nitrogen (N 2 ) to ammonium (NH 4 + ); without the bacteroids, the plant is unable to utilize N 2. The infected root cell provides some factors essential for nitrogen fixation, including leghemoglobin; this heme protein has a very high binding affinity for oxygen, which strongly inhibits nitrogenase. nucleus.")
7
Assimilation and Production of Nitrogen Compounds Most organisms can readily assimilate nitrogen from ammonia (NH 3 ) Soil bacteria quickly oxidize ammonia into nitrite (NO 2 - ) and nitrate (NO 3 - ) (nitrification reaction) Plants and microorganisms can assimilate nitrogen from nitrite and nitrate Some bacteria can use nitrate as the ultimate electron acceptor when building a proton gradient for ATP synthesis (denitrification reaction) Few bacteria can convert ammonia and nitrite into molecular nitrogen (anammox)
 Soil bacteria quickly oxidize ammonia into nitrite (NO 2 - ) and nitrate (NO 3 - ) (nitrification reaction) Plants and microorganisms can assimilate nitrogen from nitrite and nitrate Some bacteria can use nitrate as the ultimate electron acceptor when building a proton gradient for ATP synthesis (denitrification reaction) Few bacteria can convert ammonia and nitrite into molecular nitrogen (anammox)")
8
The nitrogen cycle. The total amount of nitrogen fixed annually in the biosphere exceeds 1011 kg. Reactions with red arrows occur largely or entirely in anaerobic environments. The redox states of the various nitrogen species are depicted at the bottom of the figure.

9
The anammox reactions. Ammonia and ammonium hydroxide are converted to hydrazine and H 2 O by hydrazine hydrolase, and the hydrazine is oxidized by hydrazine-oxidizing enzyme, generating N 2 and protons. The protons generate a proton gradient for ATP synthesis. On the anammoxosome exterior, protons are used by the nitrite-reducing enzyme, producing ammonium oxide and completing the cycle. All of the anammox enzymes are embedded in the anammoxosome membrane.

10
Ladderane Lipids of Anammoxosome Membrane These lipids form extremely impermeable bilayers and preventing leakage of toxic hydrazine into cytoplasm

11
Ladderane lipids of the anammoxosome membrane. The mechanism for synthesis of the unstable fused cyclobutane ring structures is unknown. (b) Ladderanes can stack to form a very dense, impermeable, hydrophobic membrane structure, allowing sequestration of the hydrazine produced in the anammox reactions.
 Ladderanes can stack to form a very dense, impermeable, hydrophobic membrane structure, allowing sequestration of the hydrazine produced in the anammox reactions..")
12
Nitrogen Fixation The reaction N 2 + 3 H 2 = 2 NH 3 –is exothermic ( H 0 = -92.4 kJ/mol) –and exergonic ( G 0 = -33.5 kJ/mol) –but very slow due to the inertness of nitrogen Only few prokaryotes produce enzymes in the nitrogenase complex that are needed for nitrogen fixation –Dinitrogenase reductase –Dinitrogenase Dinitrogenase passes electrons to N 2 and catalyzes a step-wise reduction of N 2 to NH 3 N 2 + 8 H + + 8 e - + nATP = 2 NH 3 + H 2 + nADP + nP i 2 NH 3 + 2 H + = 2 NH 4 + About 16 ADP molecules are consumed per one N 2
 –and exergonic ( G 0 = kJ/mol) –but very slow due to the inertness of nitrogen Only few prokaryotes produce enzymes in the nitrogenase complex that are needed for nitrogen fixation –Dinitrogenase reductase –Dinitrogenase Dinitrogenase passes electrons to N 2 and catalyzes a step-wise reduction of N 2 to NH 3 N H e - + nATP = 2 NH 3 + H 2 + nADP + nP i 2 NH H + = 2 NH 4 + About 16 ADP molecules are consumed per one N 2")
13
Oxidation of Pyruvate Provides Electrons to Nitrogenase Pyruvate passes electrons to ferredoxin or flavodoxin Ferredoxin or flavodoxin pass electrons to dinitrogenase reductase The reductase passes electrons to dinitrogenase Dinitrogenase passes electrons to nitrogen (or to protons?) to make NH 3 Formation of H 2 appears an obligatory side- reaction
 to make NH 3 Formation of H 2 appears an obligatory side- reaction")
14
Nitrogen fixation by the nitrogenase complex. Electrons are transferred from pyruvate to dinitrogenase via ferredoxin (or flavodoxin) and dinitrogenase reductase. Dinitrogenase reductase reduces dinitrogenase one electron at a time, with at least six electrons required to fix one molecule of N 2. An additional two electrons are used to reduce 2 H + to H 2 in a process that obligatorily accompanies nitrogen fixation in anaerobes, making a total of eight electrons required per N 2 molecule.
 and dinitrogenase reductase. Dinitrogenase reductase reduces dinitrogenase one electron at a time, with at least six electrons required to fix one molecule of N 2. An additional two electrons are used to reduce 2 H + to H 2 in a process that obligatorily accompanies nitrogen fixation in anaerobes, making a total of eight electrons required per N 2 molecule..")
15
Nitrogen Fixation Requires ATP Hydrolysis Even though thermodynamically favorable, the biological synthesis of NH 3 requires hydrolysis of large quantities of ATP Energy of ATP hydrolysis is used indirectly –Binding energy from interaction of ATP with dinitrogenase reductase increases the efficiency of reduction of dinitrogenase –ATP hydrolysis in the reductase is coupled to the reduction of dinitrogenase enzyme In addition, it is likely that ATP hydrolysis supplies some of the protons needed in the ammonia synthesis ATP 4- + H 2 O = ADP 3- + HPO 4 2- + H +

16
Dinitrogenase Complex Dinitrogenase reductase with bound ATP Dinitrogenase subunits with 4Fe-4S complexes

17
Enzymes and cofactors of the nitrogenase complex. In this ribbon diagram, the dinitrogenase subunits are shown in gray and pink, the dinitrogenase reductase subunits in blue and green. The bound ADP is red. Note the 4Fe-4S complex and the iron-molybdenum cofactor (Mo black, homocitrate light gray). The P clusters (bridged pairs of 4Fe-4S complexes) are also shown.
. The P clusters (bridged pairs of 4Fe-4S complexes) are also shown..")
18
The Iron-Molybdenum Cofactor Consists of –Seven iron atoms –Nine sulfur atoms –One molybdenum atom –One bound homocitrate The nitrogen binds to the center of the Mo-FeS cage and is coordinated to the molybdenum atom Electrons are passed to the molybdenum-bound nitrogen via the iron-sulfur complex

19
The iron-molybdenum cofactor contains 1 Mo (black), 7 Fe 9 S 1 homocitrate.
, 7 Fe 9 S 1 homocitrate.")
20
Redox Reactions in Dinitrogenase The net reaction of nitrogenase complex can be written N 2 + 8 H + + 8 e - + 16 ATP = 2 NH 3 + H 2 + 16 ADP + 16 P i Dinitrogenase reductase catalyzes –transfer of 8-electrons to dinitrogenase –hydrolysis of ATP with release of protons Dinitrogenase catalyzes –transfer of 6 electrons to nitrogen: formation of NH 3 –transfer of 2 electrons to protons: formation of H 2

21
The Mechanism of Dinitrogenase Remains Poorly Understood Extremely complex redox reaction that involves several metal atoms as cofactors and/or electron transporters A likely mechanism involves a series of single electron transfers to nitrogen

22
Broader Impact of Understanding the Nitrogen Fixation Industrial synthesis of ammonia via the Haber process is one of the most significant chemical processes for mankind –Yields over 100 million tons of fertilizer annually –This sustains life of over third of human population on Earth –Consumes non-renewable energy (1-2 % of total annual energy) –Requires high temperatures and high pressures Biological nitrogen fixation occurs at much milder conditions than the 100-year-old Haber process Biomimetic nitrogen fixation may yield significant energy savings, or allow use of renewable energy sources Understanding the biological nitrogen fixation may help in designing more efficient industrial processes
 –Requires high temperatures and high pressures Biological nitrogen fixation occurs at much milder conditions than the 100-year-old Haber process Biomimetic nitrogen fixation may yield significant energy savings, or allow use of renewable energy sources Understanding the biological nitrogen fixation may help in designing more efficient industrial processes")
23
Ammonia is Incorporated into Glutamine The glutamine amide is the source of transferable nitrogen Glutamine is made by glutamine synthetase in a two-step process Phosphorylation of glutamine creates a good leaving group that can be easily displaced by ammonia

24
Subunit structure of glutamine synthetase as determined by x-ray diffraction. The 12 subunits are identical; they are differently colored to illustrate packing and placement.

25
GS: Top view, showing active sites (green).
.")
26
Allosteric regulation of glutamine synthetase. The enzyme undergoes cumulative regulation by six end products of glutamine metabolism. Alanine and glycine probably serve as indicators of the general status of amino acid metabolism in the cell.

27
Second level of regulation of glutamine synthetase: covalent modifications. (a) An adenylylated Tyr residue
 An adenylylated Tyr residue.")
28
Cascade leading to adenylylation (inactivation) of glutamine synthetase. AT represents adenylyltransferase; UT, uridylyltransferase.

29
Proposed mechanism for glutamine amidotransferases. Each enzyme has two domains. The glutamine-binding domain contains structural elements conserved among many of these enzymes, including a Cys residue required for activity. The NH 3 -acceptor (second-substrate) domain varies. Two types of amino acceptors are shown. X represents an activating group, typically a phosphoryl group derived from ATP, that facilitates displacement of a hydroxyl group from R — OH by NH 3.
 domain varies. Two types of amino acceptors are shown. X represents an activating group, typically a phosphoryl group derived from ATP, that facilitates displacement of a hydroxyl group from R — OH by NH 3..")
30
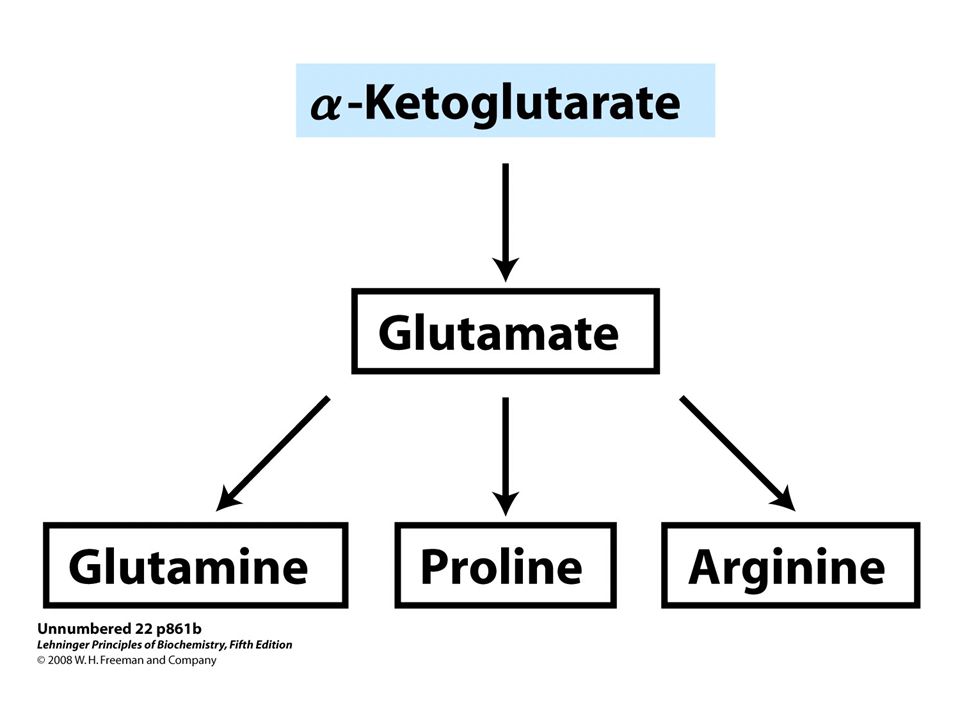
Overview of amino acid biosynthesis. The carbon skeleton precursors derive from three sources: glycolysis, the citric acid cycle and the pentose phosphate pathway.

34
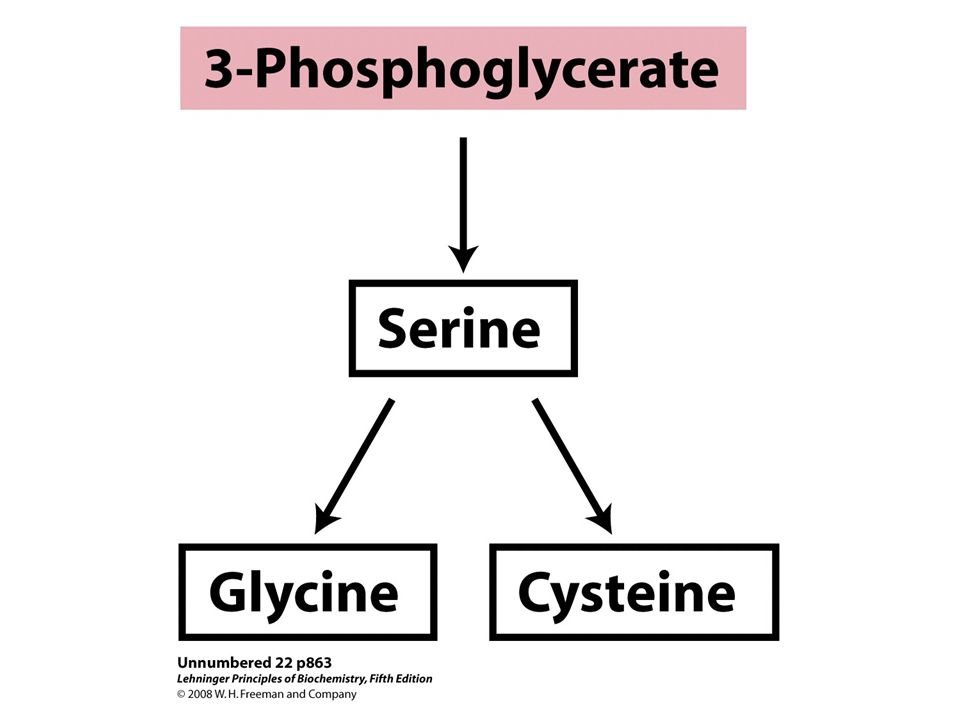
Biosynthesis of serine from 3- phosphoglycerate and of glycine from serine in all organisms. Glycine is also made from CO 2 and NH 4 + by the action of glycine synthase, with N5,N10- methylenetetrahydrofolate as methyl group donor

35
Biosynthesis of cysteine from serine in bacteria and plants. The origin of reduced sulfur is shown in the pathway on the right.

36
Biosynthesis of cysteine from homocysteine and serine in mammals. The homocysteine is formed from methionine.

38
Nucleotide Biosynthesis Nucleotides can be synthesized de novo from amino acids, ribose-5-phosphate, CO 2, and NH 3 Nucleobases can be salvaged and reused to make nucleotides Many parasites (e.g. malaria) lack de novo biosynthesis pathways and rely exclusively on salvage –Compounds that inhibit salvage pathways are promising anti-parasite drugs
 lack de novo biosynthesis pathways and rely exclusively on salvage –Compounds that inhibit salvage pathways are promising anti-parasite drugs.")
39
De Novo Biosynthesis of Purines

40
Origin of the ring atoms of purines. This information was obtained from isotopic experiments with 14C- or 15N- labeled precursors. Formate is supplied in the form of N10- formyltetrahydrofolate.

41
PRPP: phosphoribosyl pyrophosphate: How would you synthesize this?

43
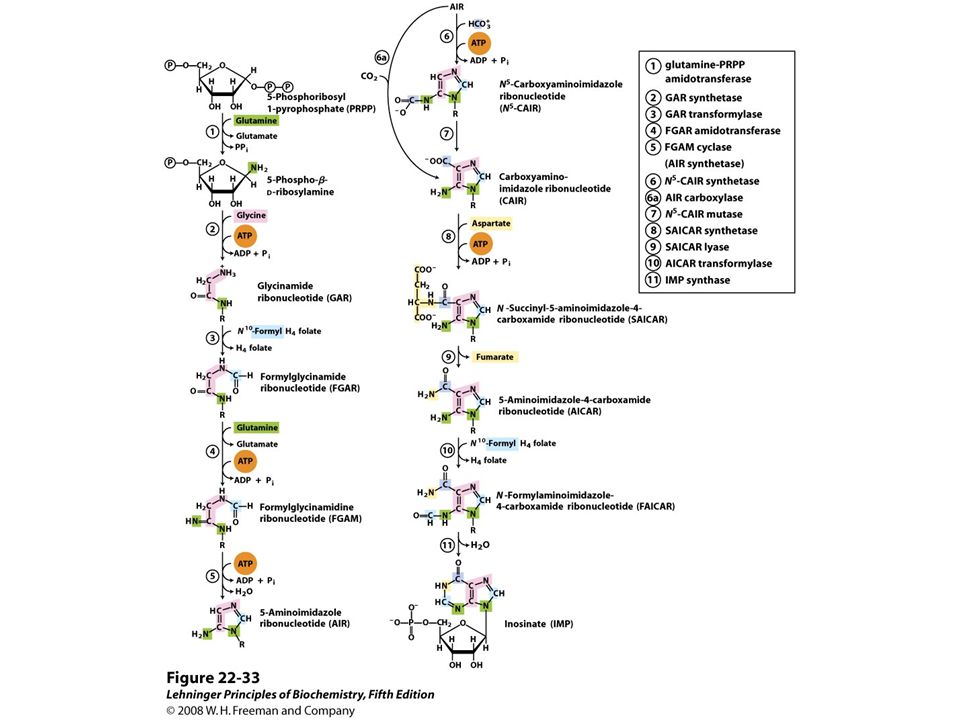
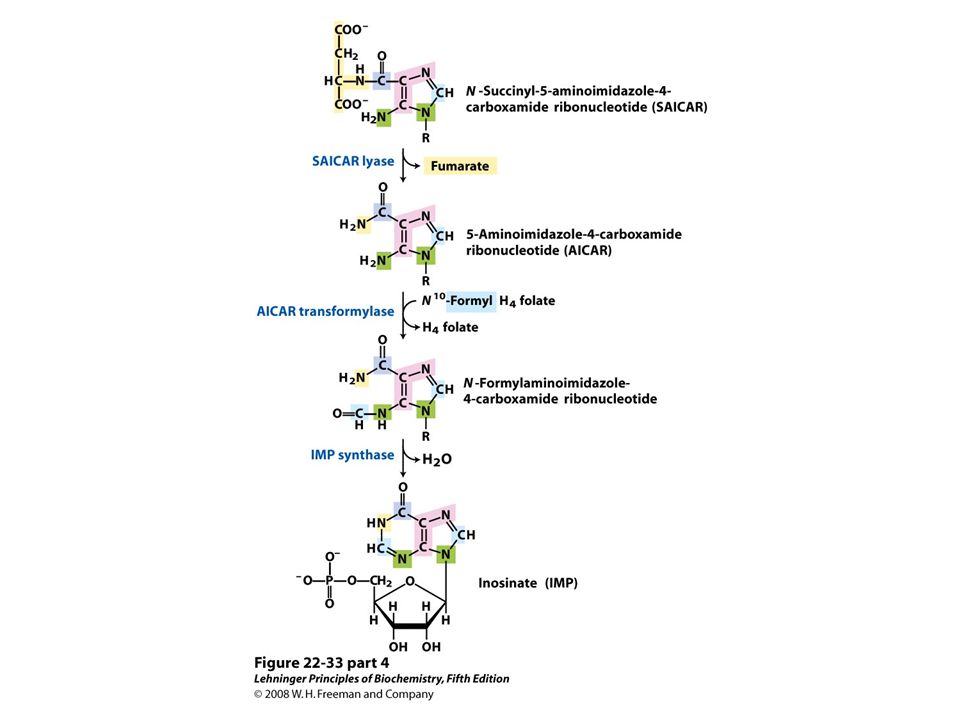
De novo synthesis of purine nucleotides: construction of the purine ring of inosinate (IMP). Formation of 5-phosphoribosylamine (step 1) is the first committed step in purine synthesis.
 is the first committed step in purine synthesis..")
44
After step 2, R symbolizes the 5- phospho-D-ribosyl group on which the purine ring is built. Formation of 5- phosphoribosylamine (step 1) is the first committed step in purine synthesis.
 is the first committed step in purine synthesis..")
45
Step 6a is the alternative path from AIR to CAIR occurring in higher eukaryotes.

47
Biosynthesis of AMP and GMP from IMP.

48
Regulatory mechanisms in the biosynthesis of adenine and guanine nucleotides in E. coli. Regulation of these pathways differs in other organisms.

49
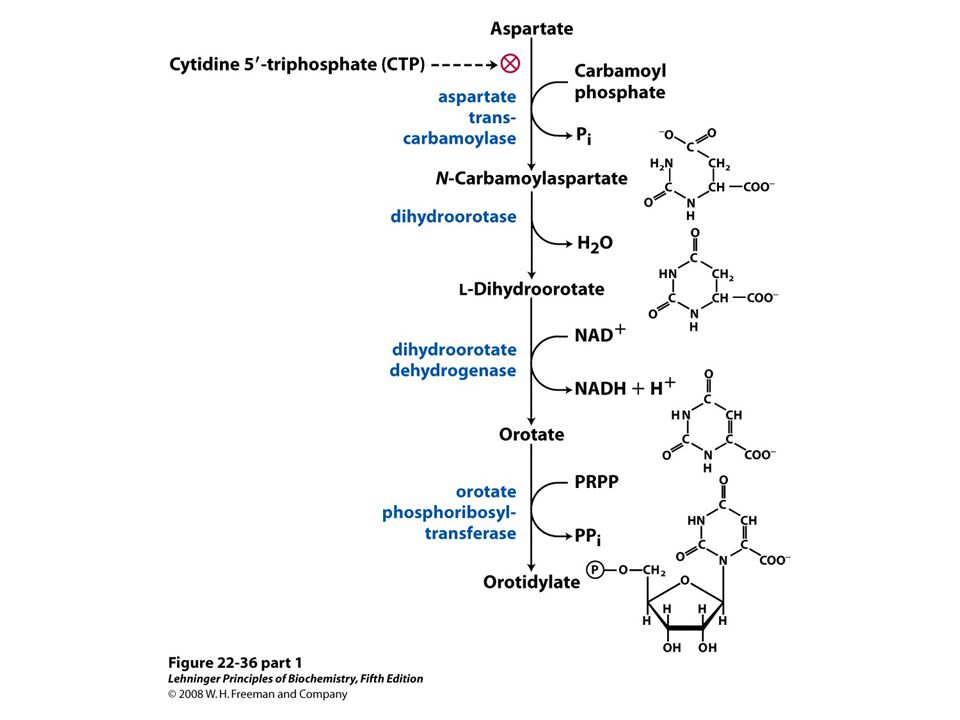
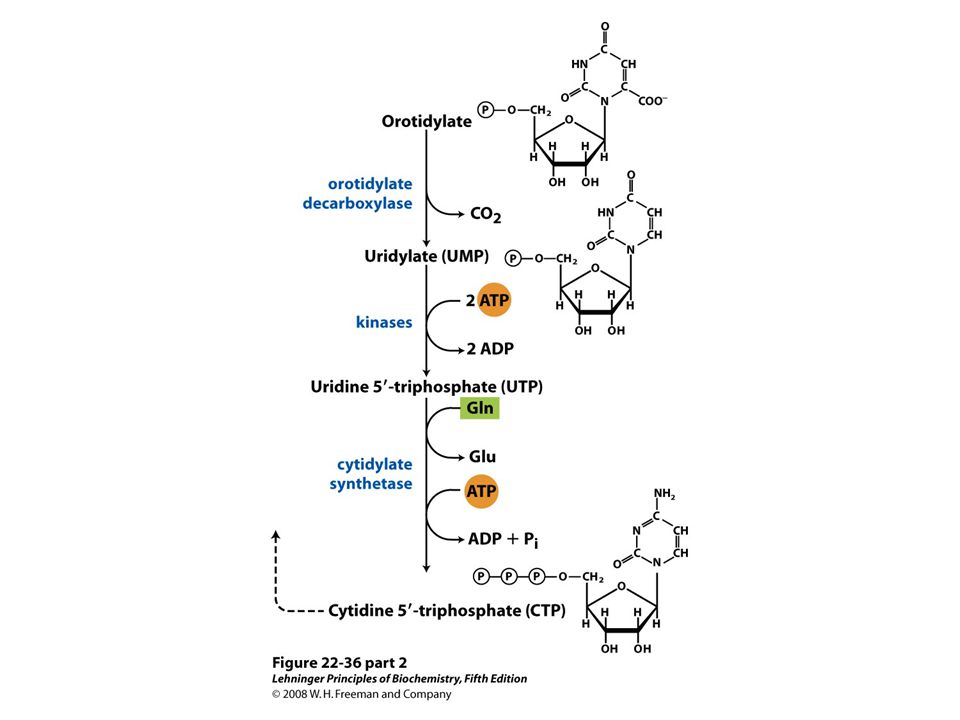
De novo synthesis of pyrimidine nucleotides: biosynthesis of UTP and CTP via orotidylate. The pyrimidine is constructed from carbamoyl phosphate and aspartate. The ribose 5-phosphate is then added to the completed pyrimidine ring by orotate phosphoribosyltransferase

52
FIGURE 22-38 Allosteric regulation of aspartate transcarbamoylase by CTP and ATP. Addition of 0.8 mM CTP, the allosteric inhibitor of ATCase, increases the K0.5 for aspartate (lower curve) and the rate of conversion of aspartate to N- carbamoylaspartate. ATP at 0.6 mM fully reverses this effect (middle curve).
 and the rate of conversion of aspartate to N- carbamoylaspartate. ATP at 0.6 mM fully reverses this effect (middle curve)..")
53
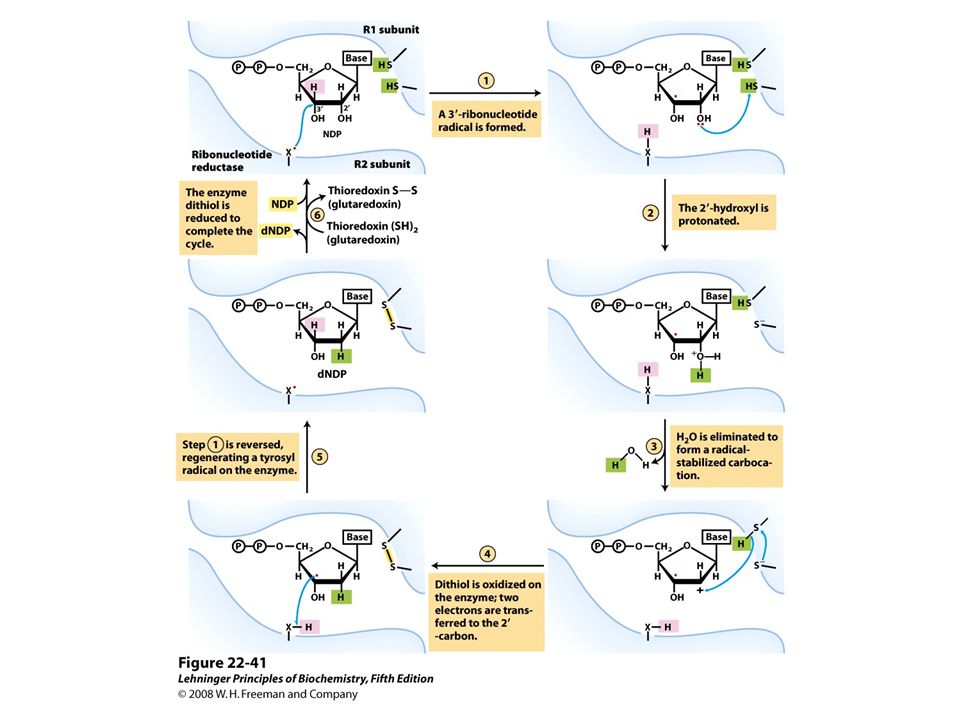
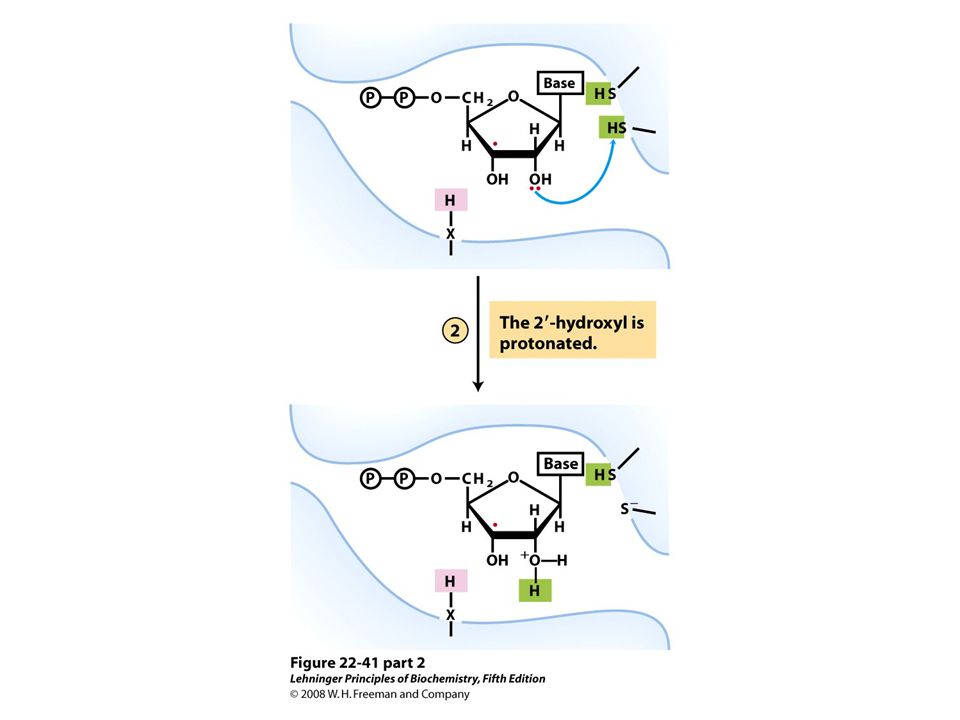
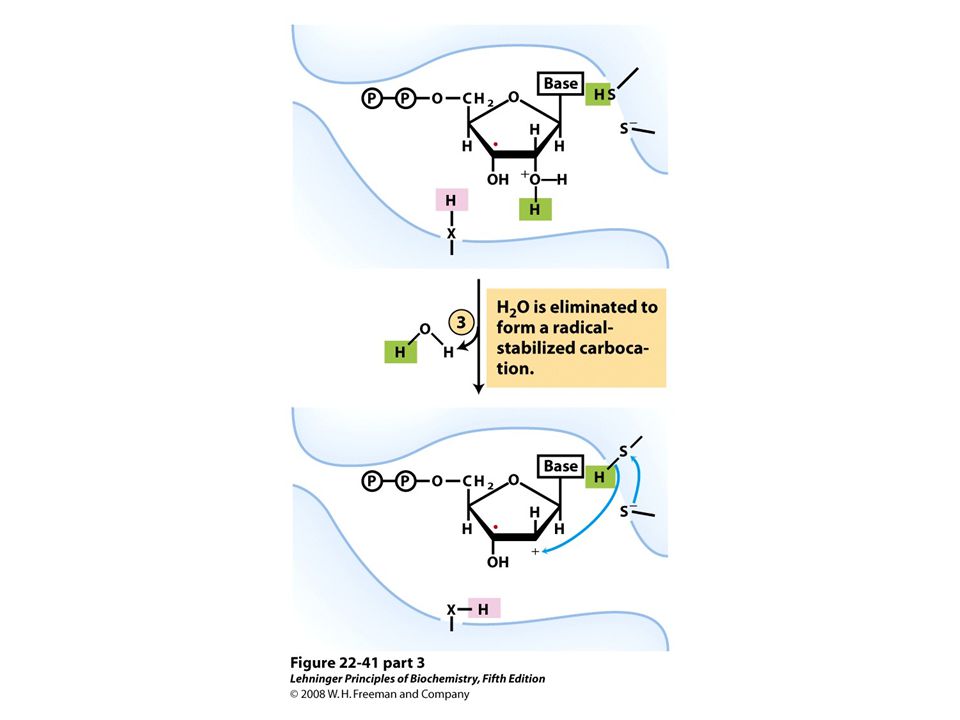
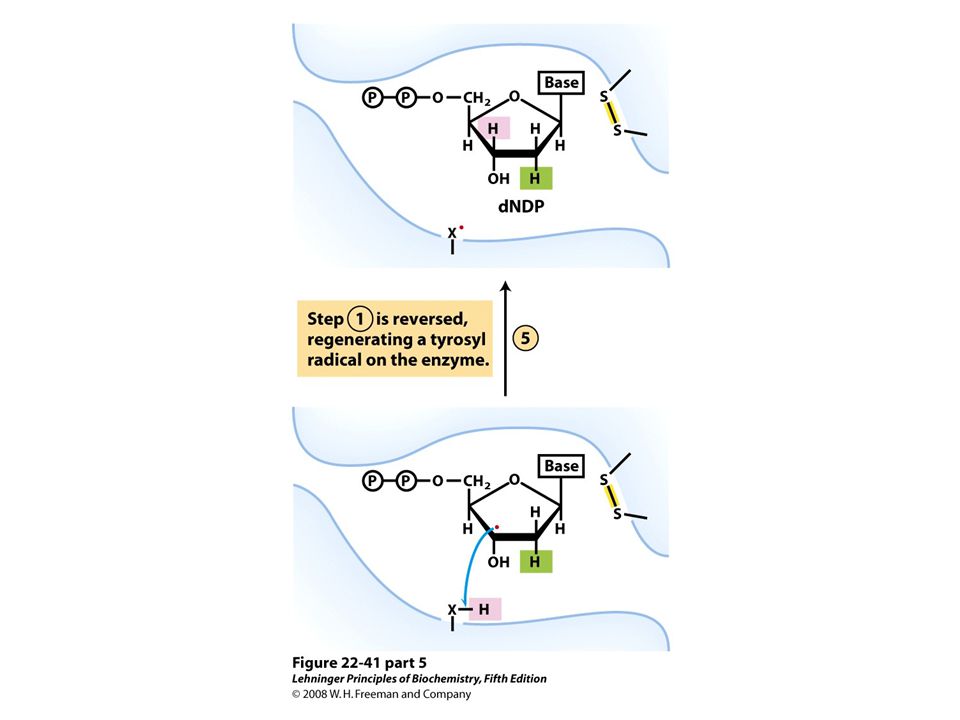
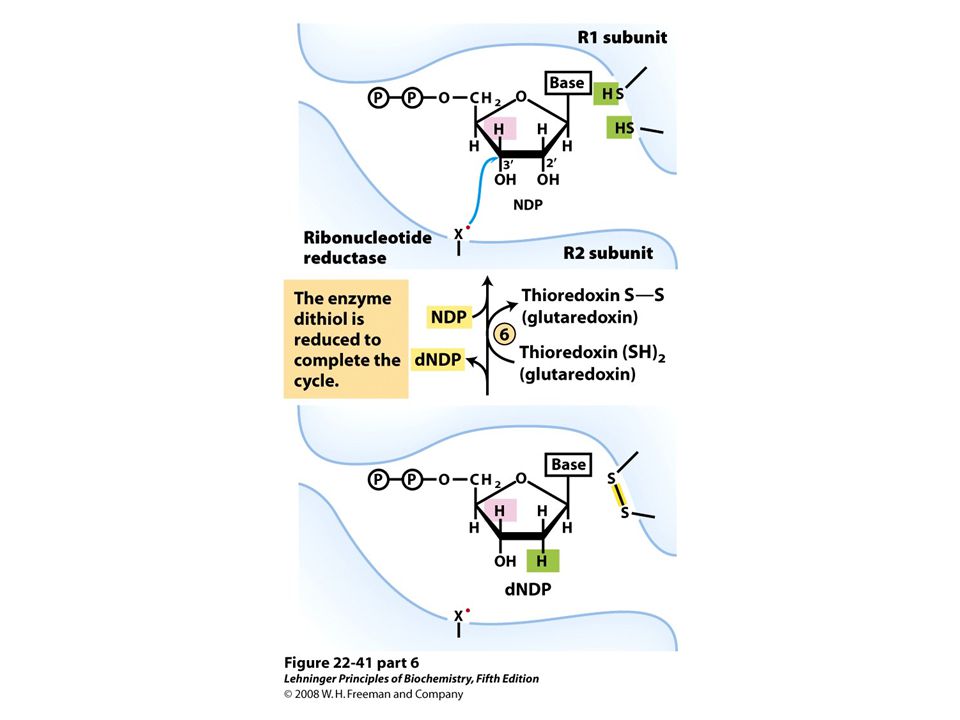
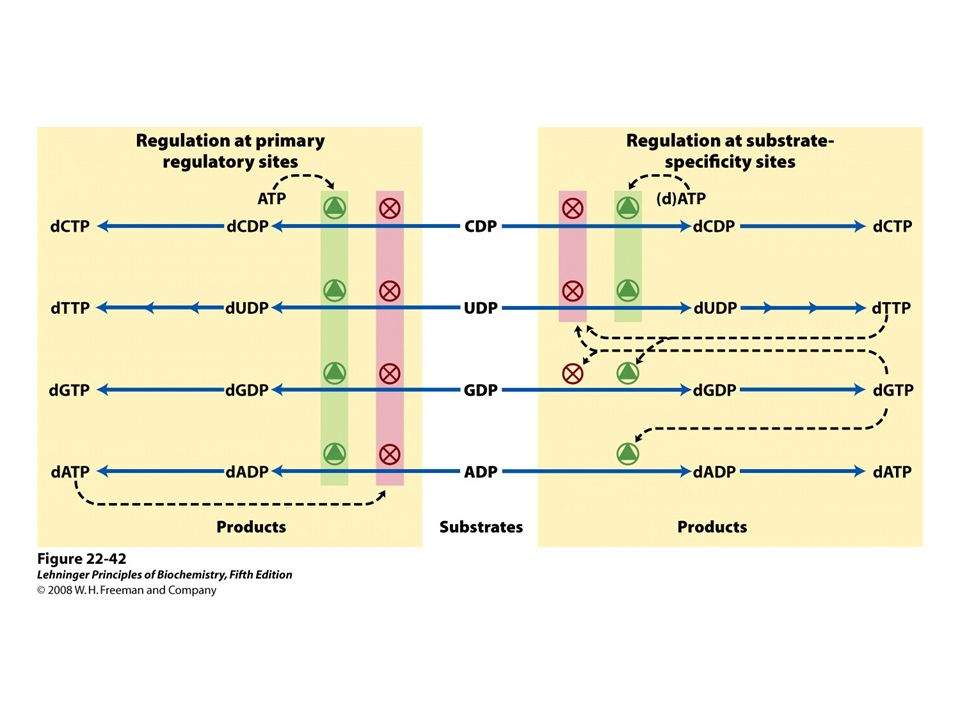
Reduction of ribonucleotides to deoxyribonucleotides by ribonucleotide reductase.

54
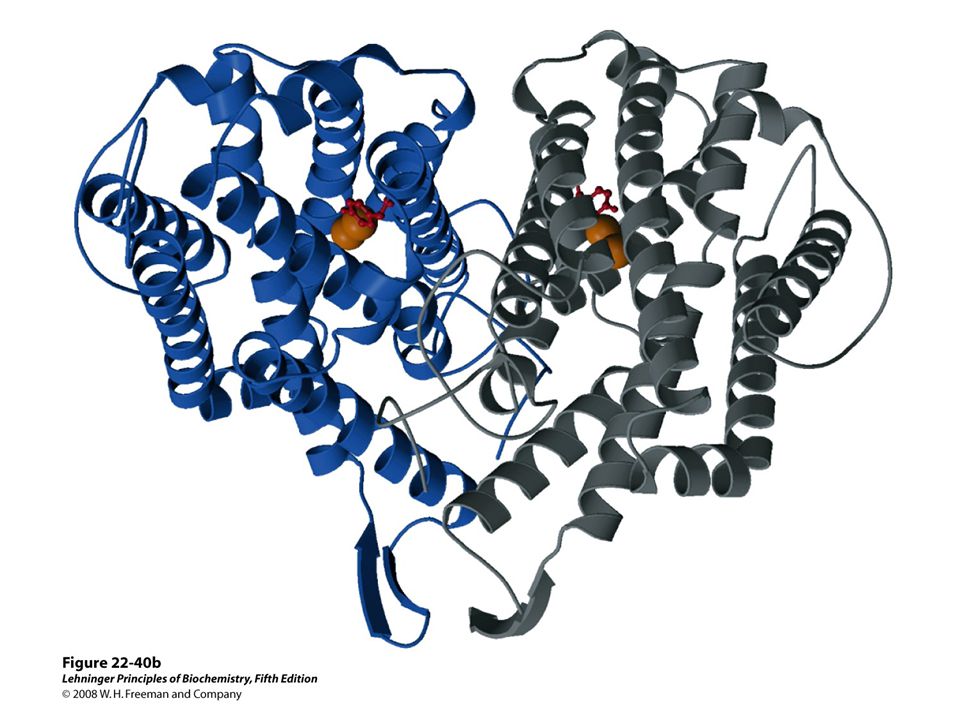
FIGURE 22-40a Ribonucleotide reductase. (a) Subunit structure. Each active site contains two thiols and a group ( — XH) that can be converted to an active-site radical; this group is probably the — SH of Cys439, which functions as a thiyl radical.
 that can be converted to an active-site radical; this group is probably the — SH of Cys439, which functions as a thiyl radical..")
56
Ribonucleotide reductase. (c) The tyrosyl radical functions to generate the active-site radical (—X ・ ), which is used in the mechanism shown in Figure 22-41.
 The tyrosyl radical functions to generate the active-site radical (—X ・ ), which is used in the mechanism shown in Figure")
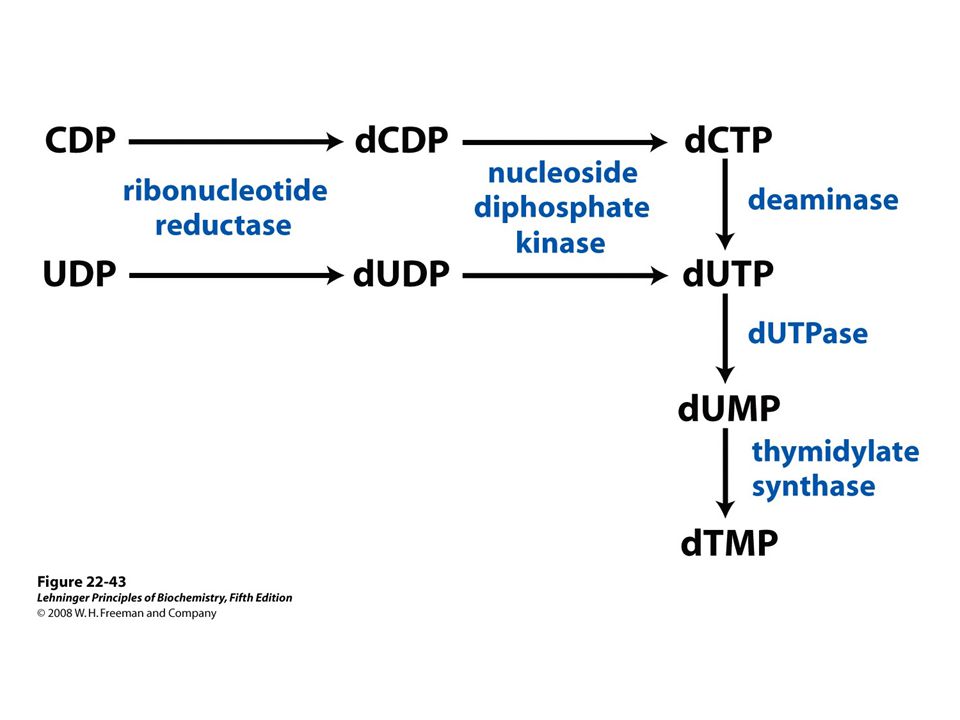
66
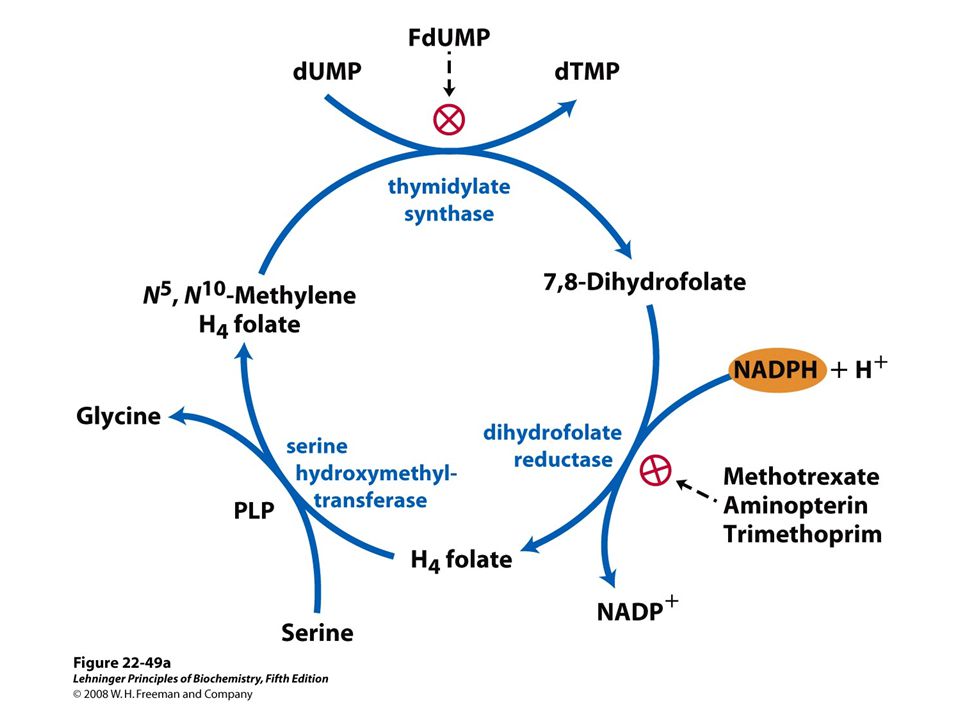
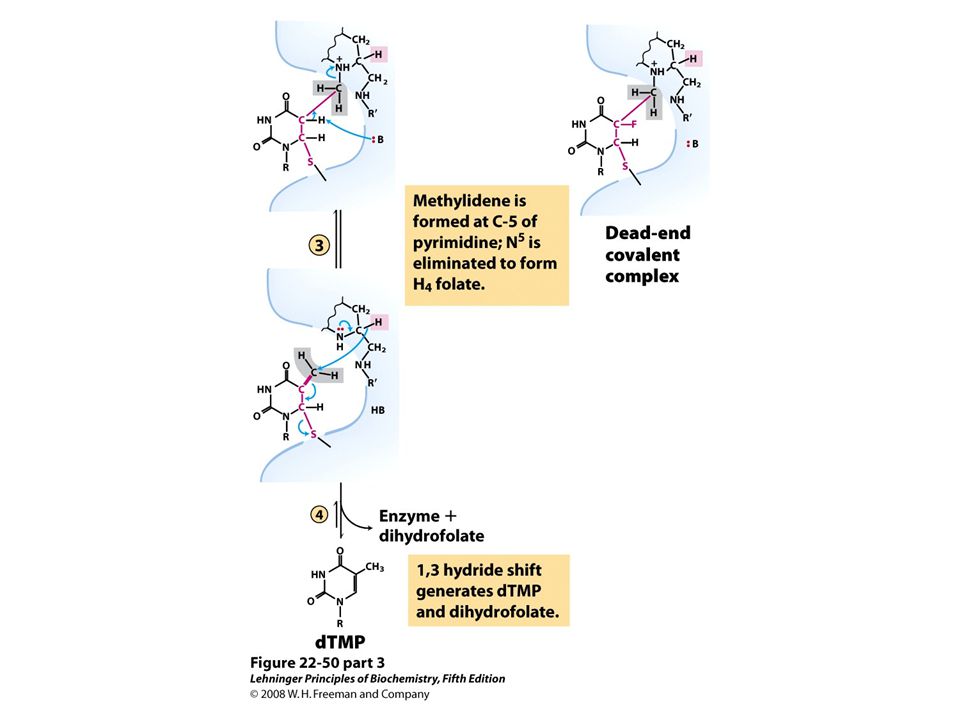
Methylation of dUMP into dTMP Thymidylate synthase is a target for some anticancer drugs

67
Aminopterin Inhibits DHF reductase and therefore dTMP synthesis Conversion of dUMP to dTMP by thymidylate synthase and dihydrofolate reductase.

69
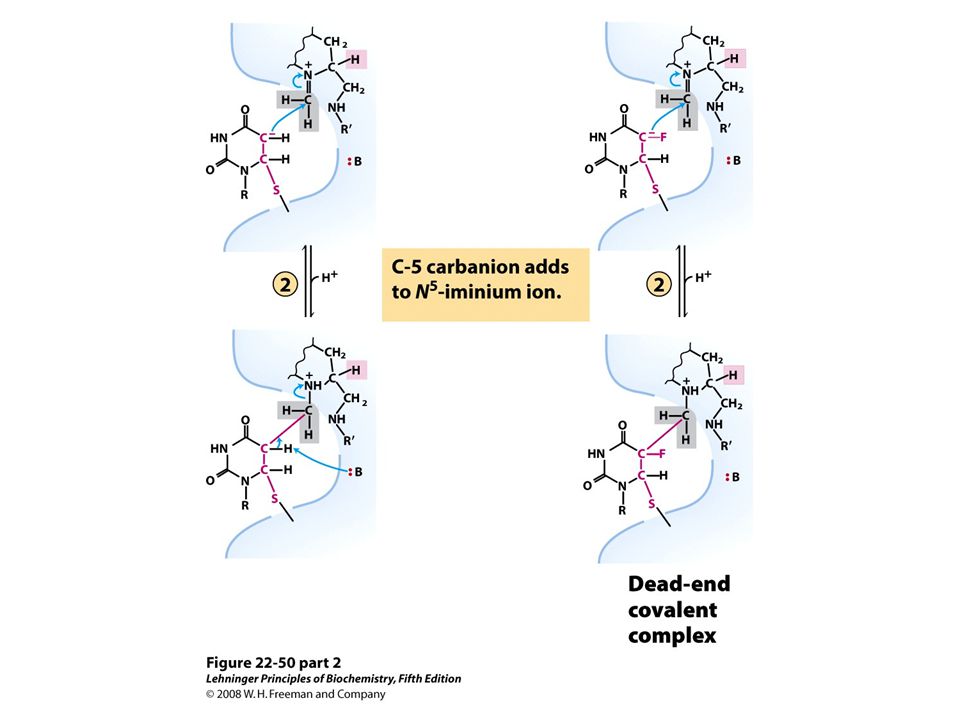
Fluorouracil and methotrexate are important chemotherapeutic agents. In cells, fluorouracil is converted to FdUMP, which inhibits thymidylate synthase. Methotrexate, a structural analog of tetrahydrofolate, inhibits dihydrofolate reductase; the shaded amino and methyl groups replace a carbonyl oxygen and a proton, respectively, in folate Another important folate analog, aminopterin, is identical to methotrexate except that it lacks the shaded methyl group. Trimethoprim, a tight- binding inhibitor of bacterial dihydrofolate reductase, was developed as an antibiotic. Thymidylate synthesis and folate metabolism as targets of chemotherapy.

70
Conversion of dUMP to dTMP and its inhibition by FdUMP

74
Nucleotide Metabolism Overview

75
HAT medium: Hypoxanthine, aminopterin, thymidine

76
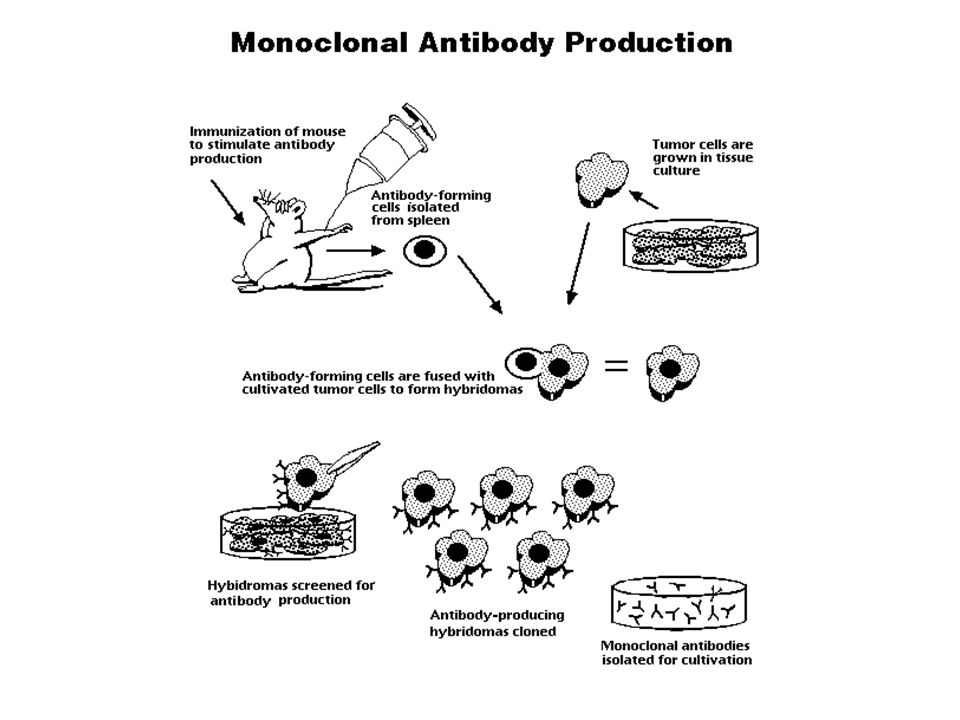
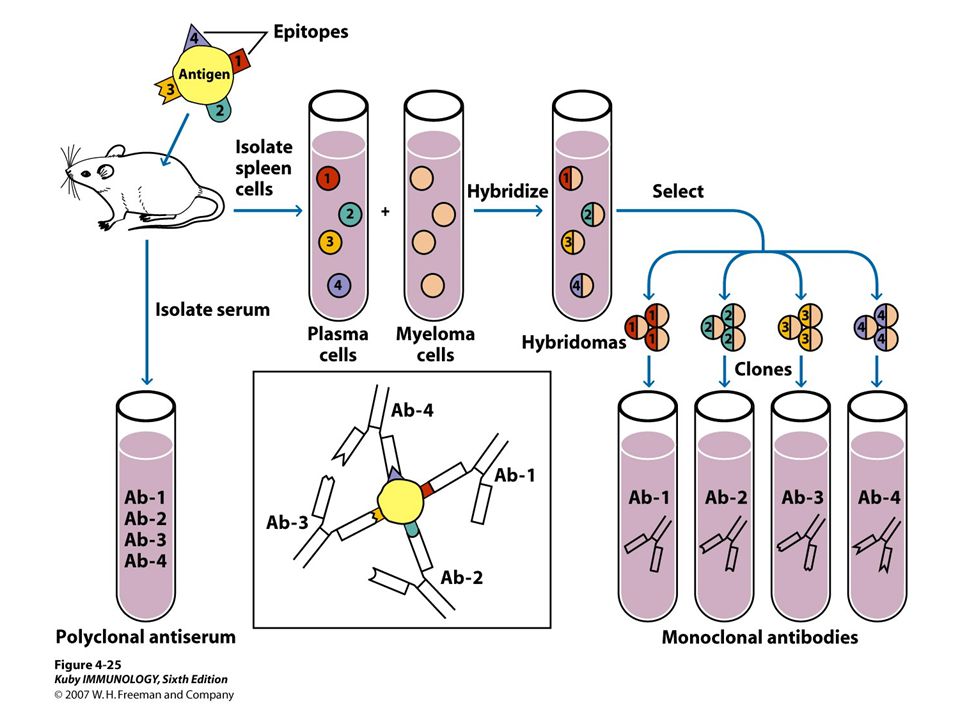
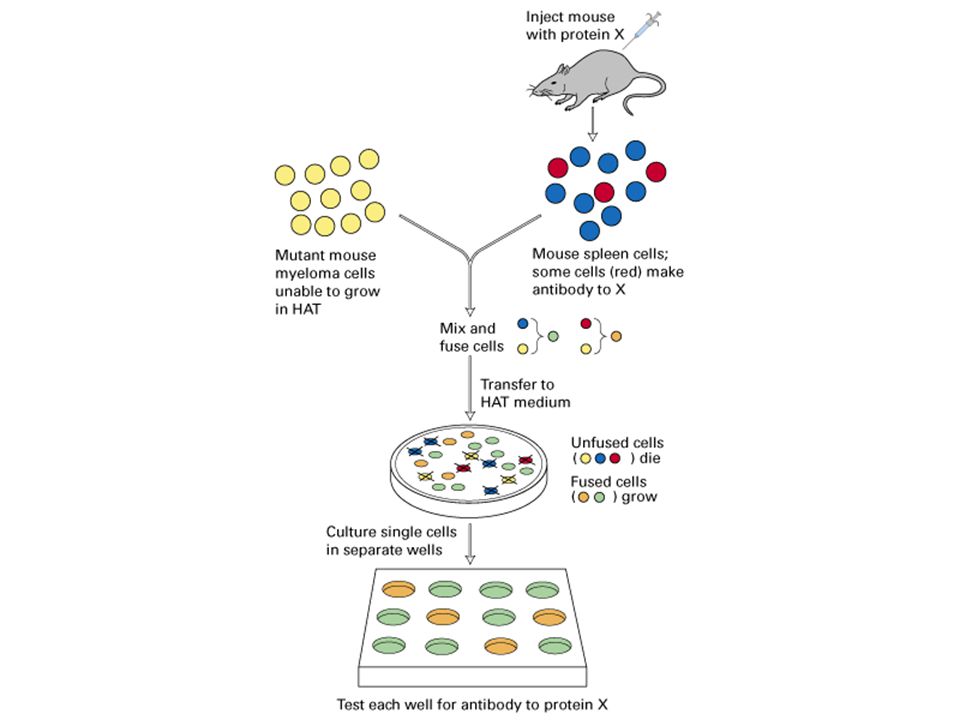
Making monoclonal antibodies

80
HAT Medium

81
“Alfonse, Biochemistry makes my head hurt!!” \

82
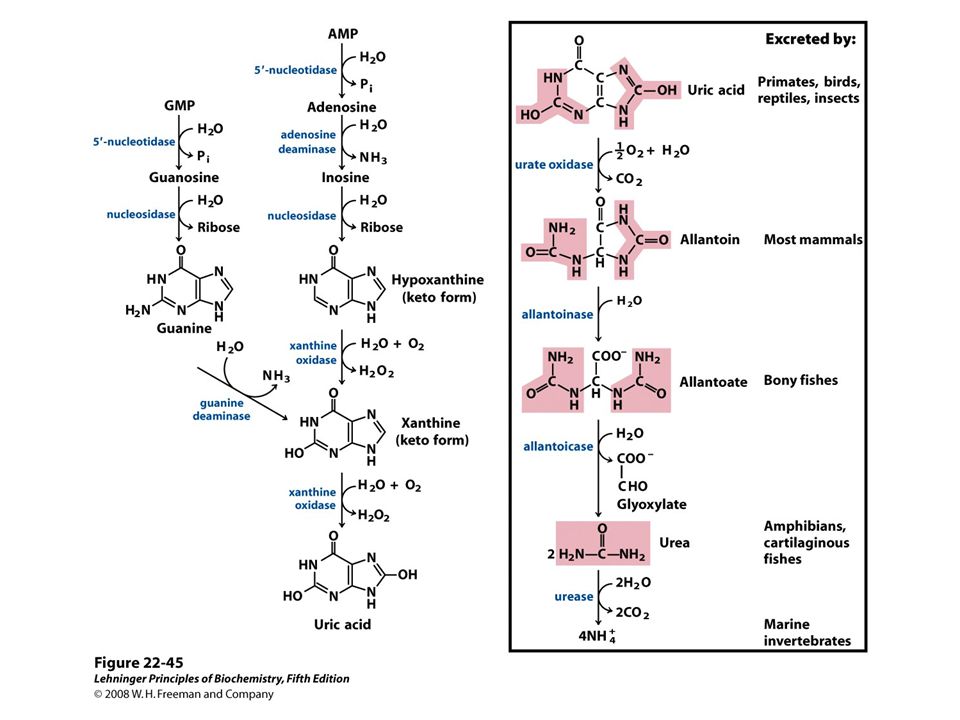
Catabolism of Purines: Formation of Uric Acid Excess purine nucleotides are dephosphorylated into nucleosides and phosphate Adenosine yields hypoxanthine via deamination and hydrolysis Guanosine yields xanthine via hydrolysis and deamination Hypoxanthine and xanthine are oxidized into urate, the anion of uric acid Spiders and other arachnids lack xanthine oxidase

84
Catabolism of Purines: Degradation of Urate to Allantoin Urate is oxidized into a 5-hydroxy- isourate by urate oxidase Hydrolysis and the subsequent decarboxylation of 5-hydroxy- isourate yields allantoin Most mammals excrete nitrogen from purines as allantoin Urate oxidase is inactive in humans and other great apes; we excrete urate Birds, most reptiles, some amphibians, and most insects also excrete urate

85
Used to reduce gout symptoms

86
Catabolism of Purines: Degradation of Allantoin Most mammals do not degrade allantoin Amphibians and fishes hydrolyze allantoin into allantoate; bony fishes excrete allantoate Amphibians and cartilaginous fishes hydrolyze allantoate into glyoxylate and urea; many excrete urea Some marine invertebrates break urea down into ammonia

87
Chapter 22: Summary Some prokaryotes are able to reduce molecular nitrogen into ammonia; understanding details of the nitrogen fixation is one of the holy grails in biochemistry The twenty common amino acids are synthesized via difficult- to-remember pathways from -ketoglutarate, 3-phospho- glycerate, oxaloacetate, pyruvate, phosphoenolpyruvate, erythrose 4-phosphate, and ribose-5-phosphate Nucleotides can be synthesized either de novo from simple precursors, or reassembled from scavenged nucleobases Purine degradation pathway in most organisms leads to uric acid but the fate of uric acid is species-specific In this chapter, we learned that:

Similar presentations










 Amino Acid Metabolism: Nitrogen Assimilation and Amino Acid Biosynthesis.>")




 Know the general mechanism of serine proteases – what imparts specificity? – how is the substrate stabilized? – how is.>")
 ATP, are the sources of.>")







