Download presentation
Presentation is loading. Please wait.

1
I corsi vengono integrati e conterranno grosso modo due moduli:
SB: systems biology ML: machine learning Alternativa 1 Alternativa 2 Mattina (Fariselli) Pomeriggio (Martelli) Lun 1 Ott SVM (ML) Grafi (SB) Mar 2 Ott System Biology (SB) Probabilita (ML) Mer 3 Ott HMM (ML) Ven 5 Ott Lun 8 Ott Mar 9 Ott Linux Linux e python Mer 10 Ott Esercitazione su SVM Gio 11 Ott Esercitazione su HMM
 Pomeriggio (Martelli) Lun 1 Ott. SVM (ML) Grafi (SB) Mar 2 Ott. System Biology (SB) Probabilita (ML) Mer 3 Ott. HMM (ML) Ven 5 Ott. Lun 8 Ott. Mar 9 Ott. Linux. Linux e python. Mer 10 Ott. Esercitazione su SVM. Gio 11 Ott. Esercitazione su HMM.")
2
Systems Biology. What Is It?
A branch of science that seeks to integrate different levels of information to understand how biological systems function. L. Hood: “Systems biology defines and analyses the interrelationships of all of the elements in a functioning system in order to understand how the system works.” It is not (only) the number and properties of system elements but their relations!!
 the number and properties of system elements but their relations!!")
3
More on Systems Biology
Essence of living systems is flow of mass, energy, and information in space and time. The flow occurs along specific networks Flow of mass and energy (metabolic networks) Flow of information involving DNA (transcriptional regulation networks) Flow of information not involving DNA (signaling networks) The Goal of Systems Biology: To understand the flow of mass, energy, and information in living systems.
 Flow of information involving DNA (transcriptional. regulation networks) Flow of information not involving DNA (signaling networks) The Goal of Systems Biology: To understand the flow of mass, energy, and information in living systems.")
4
Networks and the Core Concepts of Systems Biology
Complexity emerges at all levels of the hierarchy of life System properties emerge from interactions of components (iii) The whole is more than the sum of the parts. (iv) Applied mathematics provides approaches to modeling biological systems.
 The whole is more than the sum of the parts. (iv) Applied mathematics provides approaches to. modeling biological systems.")
5
How to Describe a System
As a Whole? Networks - The Language of Complex Systems

6
Air Transportation Network

7
The World Wide Web

8
Fragment of a Social Network
(Melburn, 2004) Friendship among 450 people in Canberra
 Friendship among 450 people in Canberra.")
9
Biological Networks A. Intra-Cellular Networks
Protein interaction networks Metabolic Networks Signaling Networks Gene Regulatory Networks Composite networks Networks of Modules, Functional Networks Disease networks B. Inter-Cellular Networks Neural Networks C. Organ and Tissue Networks D. Ecological Networks E. Evolution Network

10
The Protein Interaction Network of Yeast
Yeast two hybrid Uetz et al, Nature 2000

11
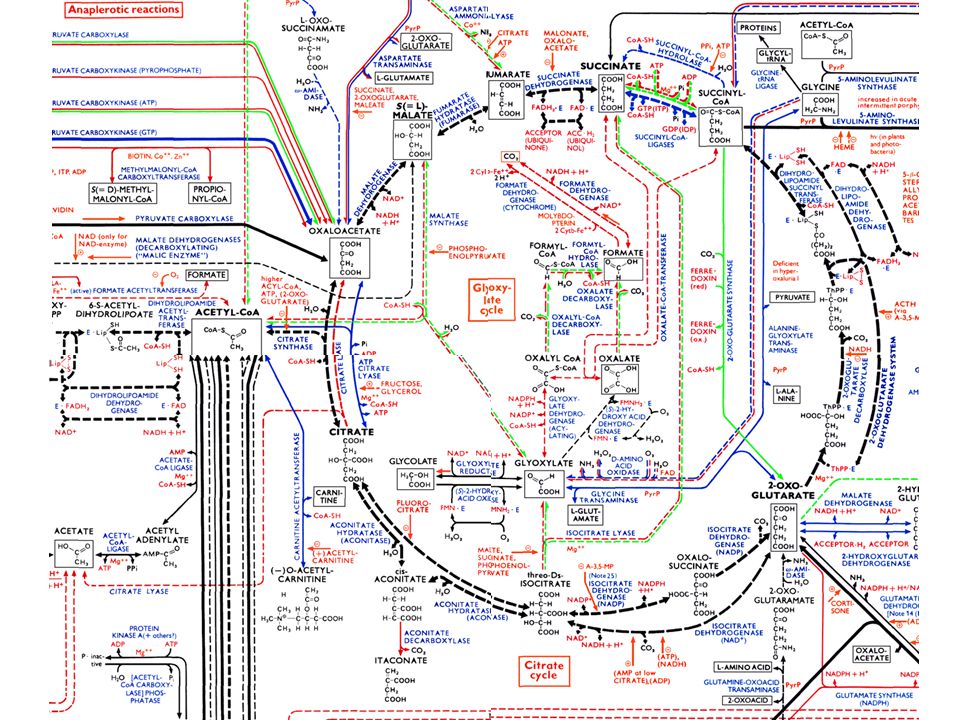
Metabolic Networks Source: ExPASy

13
Gene Regulation Networks

14
_ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _
L-A Barabasi GENOME miRNA regulation? _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ PROTEOME - - protein-gene interactions protein-protein interactions Citrate Cycle METABOLISM Bio-chemical reactions

15
Functional Networks Yeast: 1400 proteins, 232 complexes, nine functional groups of complexes Number of shared proteins 20 125 55 740 43 221 33 419 28 692 12 160 147 7 83 65 41 49 172 75 321 24 260 11 596 35 103 Cell Cycle Cell Polarity & Structure Intermediate and Energy Metabolism Protein Synthesis and Turnover Protein RNA / Transport RNA Signaling Transcription/DNA Maintenance/Chromatin Structure Number of protein complexes Number of proteins 13 111 8 61 25 40 77 19 14 97 30 16 27 299 53 37 15 22 187 73 94 Membrane Biogenesis & Turnover (Data A.-M. Gavin et al. (2002) Nature 415, ) D. Bonchev, Chemistry & Biodiversity 1(2004)
 Nature. 415, ) D. Bonchev, Chemistry & Biodiversity 1(2004)")
16
What is a Network? Network is a mathematical structure
composed of points connected by lines Network Theory <-> Graph Theory Network Graph Nodes Vertices (points) Links Edges (Lines) A network can be build for any functional system System vs. Parts = Networks vs. Nodes
 Links Edges (Lines) A network can be build for any functional system. System vs. Parts = Networks vs. Nodes.")
17
The 7 bridges of Königsberg
The question is whether it is possible to walk with a route that crosses each bridge exactly once.

18
The representation of Euler
In 1736 Leonhard Euler formulated the problem in terms of abstracted the case of Königsberg: by eliminating all features except the landmasses and the bridges connecting them; by replacing each landmass with a dot (vertex) and each bridge with a line (edge). The shape of a graph may be distorted in any way without changing the graph itself, so long as the links between nodes are unchanged. It does not matter whether the links are straight or curved, or whether one node is to the left or right of another.
 and each bridge with a line (edge). The shape of a graph may be distorted in any way without changing the graph itself, so long as the links between nodes are unchanged. It does not matter whether the links are straight or curved, or whether one node is to the left or right of another.")
19
The solution depends on the node degree
3 In a continuous path crossing the edges exactly once, each visited node requires an edge for entering and a different edge for exiting (except for the start and the end nodes). 5 3 3 A path crossing once each edge is called Eulerian path. It possible IF AND ONLY IF there are exactly two or zero nodes of odd degree. Since the graph corresponding to Königsberg has four nodes of odd degree, it cannot have an Eulerian path.
 A path crossing once each edge is called Eulerian path. It possible IF AND ONLY IF there are exactly two or zero nodes of odd degree. Since the graph corresponding to Königsberg has four nodes of odd degree, it cannot have an Eulerian path.")
20
The solution depends on the node degree
3 6 End 2 4 5 1 Start If there are two nodes of odd degree, those must be the starting and ending points of an Eulerian path.

21
Hamiltonian paths Find a path visiting each node exactly one
Conditions of existence for Hamiltonian paths are not simple

22
Hamiltonian paths

23
Graph nomenclature Graphs can be simple or multigraphs, depending on whether the interaction between two neighboring nodes is unique or can be multiple, respectively. A node can have or not self loops

24
Graph nomenclature Networks can be undirected or directed, depending on whether the interaction between two neighboring nodes proceeds in both directions or in only one of them, respectively. 1 2 3 4 5 6 The specificity of network nodes and links can be quantitatively characterized by weights 2.5 7.3 3.3 12.7 5.4 8.1 2.5 Vertex-Weighted Edge-Weighted

25
Graph nomenclature A network can be connected (presented by a single component) or disconnected (presented by several disjoint components). connected disconnected Networks having no cycles are termed trees. The more cycles thenetwork has, the more complex it is. trees cyclic graphs

26
Graph nomenclature Paths Stars Cycles Complete Graphs

27
Large graphs = Networks

28
Statistical features of networks
Vertex degree distribution (the degree of a vertex is the number of vertices connected with it via an edge)
")
29
Statistical features of networks
Clustering coefficient: the average proportion of neighbours of a vertex that are themselves neighbours Node 4 Neighbours (N) 2 Connections among the Neighbours 6 possible connections among the Neighbours (Nx(N-1)/2) Clustering for the node = 2/6 Clustering coefficient: Average over all the nodes
 2 Connections among the Neighbours. 6 possible connections among the Neighbours. (Nx(N-1)/2) Clustering for the node = 2/6. Clustering coefficient: Average over all the nodes.")
30
Statistical features of networks
Clustering coefficient: the average proportion of neighbours of a vertex that are themselves neighbours C=0 C=0 C=0 C=1

31
Statistical features of networks
Given a pair of nodes, compute the shortest path between them Average shortest distance between two vertices Diameter: maximal shortest distance How many degrees of separation are they between two random people in the world, when friendship networks are considered?

32
How to compute the shortest path between home and work?
Edge-weighted Graph The exaustive search can be too much time-consuming

33
The Dijkstra’s algorithm
Fixed nodes NON –fixed nodes Initialization: Fix the distance between “Casa” and “Casa” equal to 0 Compute the distance between “Casa” and its neighbours Set the distance between “Casa” and its NON-neighbours equal to ∞

34
The Dijkstra’s algorithm
Fixed nodes NON –fixed nodes Iteration (1): Search the node with the minimum distance among the NON-fixed nodes and Fix its distance, memorizing the incoming direction
: Search the node with the minimum distance among the NON-fixed nodes and Fix its distance, memorizing the incoming direction.")
35
The Dijkstra’s algorithm
4 Iteration (2): Update the distance of NON-fixed nodes, starting from the fixed distances Fixed nodes NON –fixed nodes
: Update the distance of NON-fixed nodes, starting from the fixed distances. Fixed nodes. NON –fixed nodes.")
36
The Dijkstra’s algorithm
Fixed nodes NON –fixed nodes The updated distance is different from the previous one Iteration: Fix the NON-fixed nodes with minimum distance Update the distance of NON-fixed nodes, starting from the fixed distances.

37
The Dijkstra’s algorithm
Fixed nodes NON –fixed nodes Iteration: Fix the NON-fixed nodes with minimum distance Update the distance of NON-fixed nodes, starting from the fixed distances.

38
The Dijkstra’s algorithm
Fixed nodes NON –fixed nodes Iteration: Fix the NON-fixed nodes with minimum distance Update the distance of NON-fixed nodes, starting from the fixed distances.

39
The Dijkstra’s algorithm
Fixed nodes NON –fixed nodes Iteration: Fix the NON-fixed nodes with minimum distance Update the distance of NON-fixed nodes, starting from the fixed distances.

40
The Dijkstra’s algorithm
Fixed nodes NON –fixed nodes Iteration: Fix the NON-fixed nodes with minimum distance Update the distance of NON-fixed nodes, starting from the fixed distances.

41
The Dijkstra’s algorithm
Fixed nodes NON –fixed nodes Conclusion: The label of each node represents the minimal distance from the starting node The minimal path can be reconstructed with a back-tracing procedure

42
Statistical features of networks
Vertex degree distribution Clustering coefficient Average shortest distance between two vertices Diameter: maximal shortest distance

43
Two reference models for networks
Regular network (lattice) Random network (Erdös+Renyi, 1959) Regular connections Each edge is randomly set with probability p
 Random network (Erdös+Renyi, 1959) Regular connections. Each edge is randomly set with probability p.")
44
Two reference models for networks
Comparing networks with the same number of nodes (N) and edges Poisson distribution Degree distribution Exp decay Average shortest path ≈ N ≈ log (N) Average connectivity high low
 and edges. Poisson distribution. Degree distribution. Exp decay. Average shortest path. ≈ N. ≈ log (N) Average connectivity. high. low.")
45
Some examples for real networks
size vertex degree shortest path Shortest path in fitted random graph Clustering Clustering in random graph Film actors 225,226 61 3.65 2.99 0.79 MEDLINE coauthorship 1,520,251 18.1 4.6 4.91 0.43 1.8 x 10-4 E.Coli substrate graph 282 7.35 2.9 3.04 0.32 0.026 C.Elegans neuron network 14 2.65 2.25 0.28 0.05 Real networks are not regular (low shortest path) Real networks are not random (high clustering)
 Real networks are not random (high clustering)")
46
Adding randomness in a regular network
Random changes in edges OR Addition of random links

47
Adding randomness in a regular network
(rewiring) Networks with high clustering (like regular ones) and low path length (like random ones) can be obtained: SMALL WORLD NETWORKS (Strogatz and Watts, 1999)
 Networks with high clustering (like regular ones) and low path length (like random ones) can be obtained: SMALL WORLD NETWORKS (Strogatz and Watts, 1999)")
48
Small World Networks A small amount of random shortcuts can decrease the path length, still maintaining a high clustering: this model “explains” the 6-degrees of separations in human friendship network

49
What about the degree distribution in real networks?
Both random and small world models predict an approximate Poisson distribution: most of the values are near the mean; Exponential decay when k gets higher: P(k) ≈ e-k, for large k.
 ≈ e-k, for large k.")
50
What about the degree distribution in real networks?
In 1999, modelling the WWW (pages: nodes; link: edges), Barabasi and Albert discover a slower than exponential decay: P(k) ≈ k-a with 2 < a < 3, for large k
, Barabasi and Albert discover a slower than exponential decay: P(k) ≈ k-a with 2 < a < 3, for large k.")
51
Scale-free networks Networks that are characterized by a power-law degree distribution are highly non-uniform: most of the nodes have only a few links. A few nodes with a very large number of links, which are often called hubs, hold these nodes together. Networks with a power degree distribution are called scale-free hubs It is the same distribution of wealth following Pareto’s law: Few people (20%) possess most of the wealth (80%), most of the people (80%) possess the rest (20%)
 possess most of the wealth (80%), most of the people (80%) possess the rest (20%)")
52
Hubs Attacks to hubs can rapidly destroy the network

53
Three non biological scale-free networks
Note the log-log scale LINEAR PLOT Albert and Barabasi, Science 1999

54
How can a scale-free network emerge?
Network growth models: start with one vertex.

55
How can a scale-free network emerge?
Network growth models: new vertex attaches to existing vertices by preferential attachment: vertex tends choose vertex according to vertex degree he In economy this is called Matthew’s effect: The rich get richer This explain the Pareto’s distribution of wealth

56
How can a scale-free network emerge?
Network growth models: hubs emerge (in the WWW: new pages tend to link to existing, well linked pages)
")
57
Metabolic pathways are scale-free
Hubs are pyruvate, coenzyme A….

58
Protein interaction networks are scale-free
Degree is in some measure related to phenotypic effect upon gene knock-out Red : lethal Green: non lethal Yellow: Unknown

59
Caveat: different experiments give different results
Titz et al, Exp Review Proteomics, 2004

60
How can a scale-free network emerge?
Gene duplication (and differentiation): duplicated genes give origin to a protein that interacts with the same proteins as the original protein (and then specializes its functions)
: duplicated genes give origin to a protein that interacts with the same proteins as the original protein (and then specializes its functions)")
61
Caveat on the use of the scale-free theory
The same noisy data can be fitted in different ways A sub-net of a non-free-scale network can have a scale-free behaviour Finding a scale-free behaviour do NOT imply the growth with preferential attachment mechanism Keller, BioEssays 2006

62
Hierarchical networks
Standard free scale models have low clustering: a modular hierarchical model accounts for high clustering, low average path and scale-freeness

63
Modules Sub-graphs more represented than expected
209 bi-fan motifs found in the E.coli regulatory network

64
Summary Many complex networks in nature and technology
have common features. They differ considerably from random networks of the same size By studying network structure and dynamics, and by using comparative network analysis, one can get answers of important biological questions.

65
Fundamental Biological Questions to Answer
(i) Which interactions and groups of interactions are likely to have equivalent functions across species? (ii) Based on these similarities, can we predict new functional information about proteins and interactions that are poorly characterized? (iii) What do these relationships tell us about the evolution of proteins, networks and whole species? (iv) How to reduce the noise in biological data: Which interactions represent true binding events? False-positive interaction is unlikely to be reproduced across the interaction maps of multiple species. Fundamental Biological Questions to Answer (i) Which interactions and groups of interactions are likely to have equivalent functions across species? (ii) Based on these similarities, can we predict new functional information about proteins and interactions that are poorly characterized? (iii) What do these relationships tell us about the evolution of proteins, networks and whole species? (iv) How to reduce the noise in biological data: Which interactions represent true binding events? False-positive interaction is unlikely to be reproduced across the interaction maps of multiple species.
 Which interactions and groups of interactions are likely to have. equivalent functions across species (ii) Based on these similarities, can we predict new functional. information about proteins and interactions that are poorly. characterized (iii) What do these relationships tell us about the evolution of proteins, networks and whole species (iv) How to reduce the noise in biological data: Which interactions. represent true binding events False-positive interaction is unlikely to be reproduced across the. interaction maps of multiple species. Fundamental Biological. Questions to Answer. (i) Which interactions and groups of interactions are likely. to have equivalent functions across species (ii) Based on these similarities, can we predict new functional. information about proteins and interactions that are poorly. characterized (iii) What do these relationships tell us about the. evolution of proteins, networks and whole species (iv) How to reduce the noise in biological data: Which. interactions represent true binding events False-positive interaction is unlikely to be reproduced. across the interaction maps of multiple species.")
66
Barabasi and Oltvai (2004) Network Biology: understanding the cell’s functional organization. Nature Reviews Genetics 5: Stogatz (2001) Exploring complex networks. Nature 410: Hayes (2000) Graph theory in practice. American Scientist 88:9-13/ Mason and Verwoerd (2006) Graph theory and networks in Biology Keller (2005) Revisiting scale-free networks. BioEssays 27.10:
 Exploring complex networks. Nature 410: Hayes (2000) Graph theory in practice. American Scientist 88:9-13/ Mason and Verwoerd (2006) Graph theory and networks in Biology. Keller (2005) Revisiting scale-free networks. BioEssays 27.10:")
Similar presentations



















 >")