Download presentation
Presentation is loading. Please wait.

1
Translation

2
Translation- the synthesis of protein from an RNA template. Five stages: Preinitiation Initiation Elongation Termination Post-translational modification

3
Translation Is the Most Complicated Biological Process Known In eukaryotes, >70 ribosomal proteins 20 (more) proteins to activate aa’s 12 (more) auxiliary enzymes 100 proteins for processing 40 tRNA and rRNAs (minimum) Other specific proteins ~300 molecules involved
 proteins to activate aa’s 12 (more) auxiliary enzymes 100 proteins for processing 40 tRNA and rRNAs (minimum) Other specific proteins ~300 molecules involved")
4
Translation Importance Location: ribosomes (predominantly Cp) Highly regulated Fast: ~20 aa/sec
 Highly regulated Fast: ~20 aa/sec")
5
Functions of the Types of RNA mRNA- serves as a template code tRNA- serves as an adapter molecule rRNA- holds molecules in the correct position, protein portion also catalyze reactions

6
The Genetic Code has been Cracked Poly U codes for poly Phe

7
The Genetic Code has been Cracked Triplet Codons Code for a Single Amino Acid UUU = Phe

8
The Genetic Code has been Cracked The Code is Commaless AUGUUU Met Phe

9
The Genetic Code has been Cracked The Code is Nonoverlapping AUGUUU Met PheNot Met Cys Val

10
The Genetic Code has been Cracked The Code is Redundant 1 codon for Met, Trp all other amino acids have 2-6 triplets

11
The Genetic Code has been Cracked The Code is Degenerate UCU Ser UCC Ser UCA Ser UCG Ser “Wobble” exists in the third position

12
The Wobble Hypothesis First two bases of codon form strong h-bonds If 1st base of anticodon is C or A (i.e. last base of codon is G or U), only one:CG and AU base pairs formed If 1st base of anticodon is G or U two codons can be recognized: U to A or G; G to C or U If 1st base of anticodon is I, three codons recognized: I to A, U or C By not having to verify all three bases, speed up process
, only one:CG and AU base pairs formed If 1st base of anticodon is G or U two codons can be recognized: U to A or G; G to C or U If 1st base of anticodon is I, three codons recognized: I to A, U or C By not having to verify all three bases, speed up process.")
13
The Genetic Code has been Cracked The Code is Unambiguous UUU doesn’t code for Phe sometimes, and other times Ser

14
The Genetic Code has been Cracked The Code is Universal Not completely true Mitochondria, e.g., have some codons different Nuance is codon usage

15
The Genetic Code has been Cracked Three Reading Frames are Possible ACUGUCGCUC... One:ACUGUCGCUC... Two:CUGUCGCUC... Three:UGUCGCUC... (Four):GUCGCUC...
:GUCGCUC....")
16
The Genetic Code has been Cracked Embedded Genes are Possible Fig. 26-3 Lehninger POB 4 th Ed. Open Reading Frame- ORF a long enough sequence between start and stop to code for legitimate protein. In viral systems, there are also examples of overlapping genes.

17
The Genetic Code has been Cracked Nonsense Codons are Stop Points UAA, UAG, and UGA

18
There is a 5’UTR Shine-Dalgarno sequence (prokaryotes) ~10 nt upstream of initiation codon Consensus: 4-9 Pu 8-13 nt upstream Positions ribosome at correct start site mRNA Structure
 ~10 nt upstream of initiation codon Consensus: 4-9 Pu 8-13 nt upstream Positions ribosome at correct start site mRNA Structure")
19
All tRNA’s Have a Similar but not Identical Structure: “Cloverleaf” ~75 nt Acceptor arm- CCA-3’ T C arm Variable length extra arm ~3-~20 nt anti-codon arm D arm canonical positions Identity elements

20
rRNA Structure (E. coli Because Well- Known) Small subunit (30S) –16S RNA –21 proteins (S1-S21) Large subunit (50S) –5S and 23S RNA –36 proteins (L1-L36) Combined = 70S Eukaryote 40S + 60S = 80S
 Small subunit (30S) –16S RNA –21 proteins (S1-S21) Large subunit (50S) –5S and 23S RNA –36 proteins (L1-L36) Combined = 70S Eukaryote 40S + 60S = 80S.")
21
Components of Mammalian Ribosomes (After Harper’s Biochemistry) ComponentMassProteinRNA No.MassSizeMassBases 40S s/u1.4x10 6 ~357x10 5 18S7x10 5 1900 60S s/u2.8x10 6 ~357x10 5 18S7x10 5 1900 ~501x10 6 5S35,000120 5.8S45,000160 28S 1.6x10 6 4700
 ComponentMassProteinRNA No.MassSizeMassBases 40S s/u1.4x10 6 ~357x S7x S s/u2.8x10 6 ~357x S7x ~501x10 6 5S35, S45, S 1.6x")
22
rRNA Structure Scientific American circa 1960 and Science 2002

23
The P Site and the A Site Fig. 27-11 Lehninger POB 3 rd Ed.

24
The Preinitiation Stage is Characterized by Formation of Required Starting Complexes

25
Preinitiation - Charging the tRNA

26
Aminoacyl-tRNA Synthetase One for each amino acid 2 step mechanism: 1. attach a.a. to AMP 2a. transesterify to 3’ (class 2) 2b. transesterify to 2’ and then rearrange(class 1) Proofread –identity elements –“sieve”
 2b. transesterify to 2’ and then rearrange(class 1) Proofread –identity elements – sieve .")
27
Mechanism of Class I vs. Class II Fig. 27-14 Lehninger POB 4 th Ed.

28
Perspective on Scale Fig. 27-17(a) Lehninger POB 4 th Ed.
 Lehninger POB 4 th Ed.")
29
Preinitiation: Formylation of met- tRNA fmet (Prokaryotes Only)
")
30
Preinitiation: Dissociation of Ribosomes (IF-1 and IF-3)
")
31
Preinitiation: IF-2:GTP Binary Complex Formation IF-2B represents a guanine exchange molecule

32
Preinitiation: IF-2:GTP:Charged tRNA Ternary Complex Formation

33
Preinitiation: 40S Preinitiation Complex

34
Initiation: IF-4F, 4A and 4B Bind mRNA to Place it on Small Subunit

35
Initiation: 40S Initiation Complex

36
Initiation: 80S Initiation Complex

37
Initiation Preinitiation complexes form an 80S complex: small subunit, ternary complex (GDP + Pi leave), mRNA, large subunit, aminoacyl tRNA P-site- only thing that can enter is a peptide In prokaryotes, f-met “tricks” the ribosome A-site- only thing that can enter is an aminoacyl tRNA
, mRNA, large subunit, aminoacyl tRNA P-site- only thing that can enter is a peptide In prokaryotes, f-met tricks the ribosome A-site- only thing that can enter is an aminoacyl tRNA")
38
Elongation 1. EF-1:GTP:aminoacyl-tRNA ternary complex enters A-site; GDP + Pi leave (EF-Tu and EF-Ts involved with GTP metabolism in prokaryotes) 2. Peptide bond forms as P-site content is transferred onto A-site occupant 3. Translocation requires GTP-bound EF-2
 2. Peptide bond forms as P-site content is transferred onto A-site occupant 3. Translocation requires GTP-bound EF-2.")
39
Elongation I

40
Elongation: Peptide Bond Forms as P- site Content is Transferred Onto A-Site Occupant

41
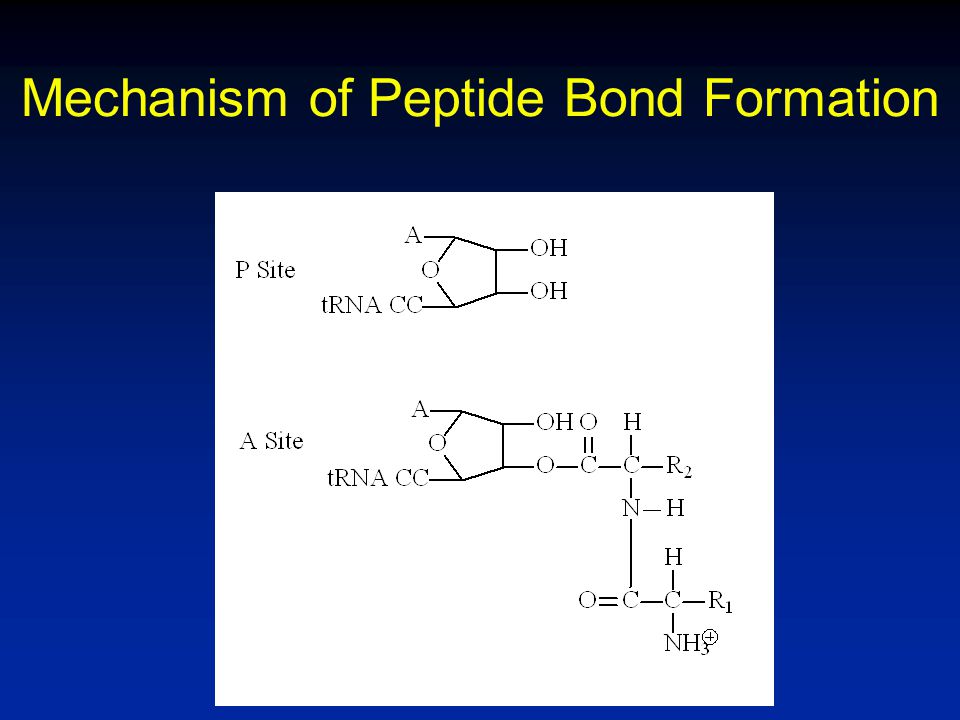
Mechanism of Peptide Bond Formation

43
Elongation: Translocation Requires GTP-bound EF-2

44
Termination 1. UAA, UAG, UGA is enveloped by A-site of ribosome 2. RF-1 enters A site 3. GTP is hydrolyzed, H 2 O is used to cleave protein off tRNA 4. Components are recycled to synthesize another protein molecule

45
Termination: UAA, UAG, or UGA is Enveloped by A-site of Ribosome

46
Termination: RF-1 Enters A Site

47
Termination: GTP is Hydrolyzed and H 2 O is Used to Cleave Protein Off tRNA

48
Energetics Each amino acid residue requires >4 ATP equivalents ATP→AMP + PPi to “charge” tRNA 1 GTP is used to place aminoacyl-tRNA into A-site 1 GTP is used to translocate after each peptide bond formation ATP hydrolysis for unknown purpose at…

49
Regulation of Translation: eEF 2 a. eEF 2 phosphorylated under stress b. when phosphorylated, eEF 2B doesn’t allow GDP-GTP exchange and protein synthesis stops

50
Regulation of Translation: eIF 4F 4F is complex of 4E (cap binding protein) and 4A (ATPase that unwinds RNA) and 4G (function unknown) a.eIF-4E can be phosphorylated: why or how unknown- but this activates b. 4E-BP complex forms which inactivates 4E. Phosphorylation in presence of insulin dissociates complex

51
Post-translational Modifications 1. Proteolytic cleavage (most common) Signal sequences; zymogens 2. Disulfide bond formation: not as well understood 3. Group addition a. Glycosylation (most complex known) b. Acetylation or phosphorylation, etc. c. Farnesyl or Geranylgeranyl 4. Amino acid modification a. Hydroxylation of Pro (in ER) b. Methylation of Lys 5. Other things
 Signal sequences; zymogens 2. Disulfide bond formation: not as well understood 3. Group addition a. Glycosylation (most complex known) b. Acetylation or phosphorylation, etc. c. Farnesyl or Geranylgeranyl 4. Amino acid modification a. Hydroxylation of Pro (in ER) b. Methylation of Lys 5. Other things.")
52
Signal Sequence: Human PreProInsulin Met Ala Leu Trp Met Arg(+) Leu Leu Pro Leu Leu Ala Leu Leu Ala Leu Trp Gly Pro Asp Pro Ala Ala Ala Phe Val
 Leu Leu Pro Leu Leu Ala Leu Leu Ala Leu Trp Gly Pro Asp Pro Ala Ala Ala Phe Val")
53
SRP Ribosome Cycle Fig. 27-33 Lehninger POB 4 th Ed.

54
Other Signal Sequences Fig. 27-39 Lehninger POB 3 rd Ed. KDEL anywhere in protein directs back to ER Chloroplast and Mitochondria: –Different in that whole protein is made and then directed via chaperone proteins –Different sequences: mitochondrial not contiguous

55
Other Signal Sequences Fig. 27-37 Lehninger POB 4 th Ed. KDEL anywhere in protein directs back to ER Chloroplast and Mitochondria: –Different in that whole protein is made and then directed via chaperone proteins –Different sequences: mitochondrial not contiguous Nuclear signal: –Different in that not cleaved

56
Other Signal Sequences KDEL anywhere in protein directs back to ER Chloroplast and Mitochondria: –Different in that whole protein is made and then directed via chaperone proteins –Different sequences: mitochondrial not contiguous Nuclear signal: –Different in that not cleaved Doesn’t have to be protein signal: M6P targets to lysosome

57
Other Proteolytic Events Met aminopeptidase Proprotein vs preproprotein

58
Disulfide Bond Formation is Not Well Understood Occurs in ER PDI and Chaperones involved PDI “shuffles” disulfide bonds Chaperones consist of two rings of identical s/u’s and somehow facilitate appropriate interactions

59
Glycosylation Occurs in Multiple Compartments (ARB figure) Oligo “tree” transferred en bloc from dolichol-based structure Some trimming takes place before export from ER Often, further trimming in cis-Golgi…
 Oligo tree transferred en bloc from dolichol-based structure Some trimming takes place before export from ER Often, further trimming in cis-Golgi…")
60
Glycosylation Continued …before building up with different sugars takes place Packaging occurs as the proteins pass through the trans- golgi network Exocytosis into bloodstream takes place Microheterogeneity

61
Lipid Moieties Fig. 27-30 Lehninger POB 4 th Ed. CaaX farnesylation signal (a for aliphatic) aaX removed after farnesyl added CXX or CXC geranylgeranylation signal (X for anything)
 aaX removed after farnesyl added CXX or CXC geranylgeranylation signal (X for anything).")
62
Protein Degradation: The Ultimate in Posttranslational Modification Regulated: 1. The N-end Rule not a sensitive system

63
The N-End Rule

64
Protein Degradation Regulated: 1. The N-end Rule not a sensitive system 2. Lysosome endocytosis 3. The Ubiquitin system

65
Ubiquination Fig. 27-41 Lehninger POB 4 th Ed. Ubiquitin common throughout the animal kingdom 76 aa (8.5 kDa) i.e. small 53% homology between yeast and humans E1 (ubiquitin activating enzyme) E2 (ubiquitin carrier protein) E3 (ubiquitin protein ligase) selects proteins to tag by N-term
 i.e. small 53% homology between yeast and humans E1 (ubiquitin activating enzyme) E2 (ubiquitin carrier protein) E3 (ubiquitin protein ligase) selects proteins to tag by N-term.")
66
More Than You Ever Wanted to Know About Ubiutin Science 315: 201-205, 2007 Monoubiquitination- a single Ub is attached Multiubiquitination- several residues of target protein each have single Ub is attached Polyubiquitination- numerous copies of Ub attached to same site of target protein. Ub has 7 different Lys, so many ways to do this. Once polyUb’d, targeted for destruction Ub can be removed from protein So clearly there must be mechanism to determine level of Ub

67
Ub is Now Known to Do More Than Just Target for Destruction Science 315: 201-205, 2007 In yeast, monoUb is sufficient as an endocytic internalization signal (less clear in animals) –necessary, sufficient When epidermal growth factor receptor (EGFR) was stimulated at low epidermal growth factor (EGF) concentrations in HeLa cells, EGFR ubiquitination was not detected, and the receptor localized with clathrin; however, at high EGF concentrations, EGFR was ubiquitinated, and the receptor localized with both caveolae and clathrin.
 –necessary, sufficient When epidermal growth factor receptor (EGFR) was stimulated at low epidermal growth factor (EGF) concentrations in HeLa cells, EGFR ubiquitination was not detected, and the receptor localized with clathrin; however, at high EGF concentrations, EGFR was ubiquitinated, and the receptor localized with both caveolae and clathrin.")
68
Ub is Now Known to Do More Than Just Target for Destruction Science 315: 201-205, 2007 Endocytosis of certain membrane proteins seems to require polyubiquitination. For example, in the case of the ß 2 AR, (recall this is a G protein–coupled receptor), agonist stimulation leads to rapid polyubiquitination of both the receptor and the receptor regulatory protein ß- arrestin....anthrax toxin triggers ubiquitination of its receptor to facilitate efficient and rapid endocytosis of the toxin- receptor complex.... important for toxin action, because passage through low-pH endosomal compartments makes the toxin competent to induce toxicity in cells Protein ubiquitination is best compared to protein phosphorylation.
, agonist stimulation leads to rapid polyubiquitination of both the receptor and the receptor regulatory protein ß- arrestin....anthrax toxin triggers ubiquitination of its receptor to facilitate efficient and rapid endocytosis of the toxin- receptor complex.... important for toxin action, because passage through low-pH endosomal compartments makes the toxin competent to induce toxicity in cells Protein ubiquitination is best compared to protein phosphorylation..")
69
Ub is Now Known to Do More Than Just Target for Destruction Science 315: 201-205, 2007

Similar presentations



 bring amino acids (AA),>")



>")

 Protein Synthesis. Translation Slow rate of synthesis (18 amino acids per second) In bacteria translation and transcription are coupled.>")









