Download presentation
Presentation is loading. Please wait.

2
Section L – Regulation of transcription in prokaryotes

3
L1 The lac operon The operon, The lactose operon, The lac repressor, Induction, cAMP receptor proteinThe operonThe lactose operonThe lac repressor InductioncAMP receptor protein L2 The trp operon The tryptophan operon, The trp repressor, The attenuator, Leader RNA structure, The leader peptide, Attenuation, Importance of attenuationThe tryptophan operonThe trp repressorThe attenuatorLeader RNA structureThe leader peptideAttenuationImportance of attenuation L3 Transcriptional regulation by alternative σ factor Sigma factor, Promoter recognition, Heat shock, Sporulation in Bacteriophage subtilis, Baxteriophage σ factorsSigma factorPromoter recognitionHeat shock Sporulation in Bacteriophage subtilis Baxteriophage σ factorsContents

4
L1 The lac operon — The operon Operon: Operon: a unit of prokarytoic gene expression which typically includes: 1. Structural genes for enzymes in a specific biosynthetic pathway whose expression is co- ordinately controlled 2. Control elements, such as operator sequence 3. Regulator gene(s) whose products recognize the control elements.
 whose products recognize the control elements..")
5
Control element Structural genes

6
The Lac Operon The Trp Operon Transcriptional regulation by alternative σ Factors The operon, the lactose operon, the lac repressor, induction, cAMP receptor protein The trp operon, the trp repressor, the attenuator, leader RNA structure, the leader peptide, attenuation & its importance Sigma factor, promoter recognition, heat shock, sporulation in B. subtilis, bacteriophage factors

7
L1 The lac operon — The lactose operon E. coli can use lactose as a source of carbon. However, the enzymes required for the use of lactose as a carbon source are only synthesized when lactose is available as the sole carbon source.

8
lacY encodes a galactoside permease to transport Lactose across the cell wall lacZ codes for β -galactosidase for lactose hydrolysis lacA encodes a thiogalactoside transacetylase for lactose metabolism

9
The lacZ, lacY, lacA genes are transcribed from a single (lacZYA) transcription unit under the control of a signal promoter P lac. LacZYA transcription unit contains an operator site O lac position between bases -5 and +21 at the 3’-end of P lac Binds with the lac repressor

10
L1 The lac operon — The lac repressor The repressor is encoded by LacI and active as a tetramer consisting of 4 identical subunits (has a symmetrical structure). It binds to occupies the operator-binding site Olac (28bp, palindromic) and blacks almost all transcription of lacZYA when lack of inducer (such as lactose).
 and blacks almost all transcription of lacZYA when lack of inducer (such as lactose)..")
11
The repressor and RNA polymerase can bind simultaneously to the lac promoter and operator sites. The lac repressor actually increases the binding of the polymerase to the lac promoter by two orders of magnitude. Thus, RNA polymerase binds very tightly to Plac but no transcription occur because of the bound repressor

12
L1 The lac operon — Induction When lac repressor binds to the inducer (whose presence is dependent on lactose), it changes conformation and cannot bind to Olac site any more. This allows rapid induction of lacZYA transcription.

13
i p o z y a Very low level of lac mRNA Absence of lactose Active i p o z y a -Galactosidase Permease Transacetylase Presence of lactose Inactiv e Lack of inducer: the lac repressor block all but a very low level of trans- cription of lacZYA. Lactose is present, the low basal level of permease allows its uptake, andβ- galactosidase catalyzes the conversion of some lactose to allolactose. Allolactose acts as an inducer, binding to the lac repressor and inactivate it.

14
Allolactose causes a change in the conformation of the repressor tetramer, reducing its affinity for the lac operator. The lac operator is removed from the Olac and allows the polymerase to rapidly begin transcription of the lacZYA. Lactose (allolactose) is a native inducer to release RNA transcription elongation from Plac. IPTG, a synthetic inducer, can rapidly simulate transcription of the lac operon structural genes. IPTG is used to induce the expression of the cloned gene from LacZ promoter in many vectors, such as pUC19.
 is a native inducer to release RNA transcription elongation from Plac. IPTG, a synthetic inducer, can rapidly simulate transcription of the lac operon structural genes. IPTG is used to induce the expression of the cloned gene from LacZ promoter in many vectors, such as pUC19..")
15
Amp r ori pUC18 (3 kb) Lac promoter lacZ’ Gene X No IPTG, little expression of X gene With IPTG, efficient expression of X gene. MCS (Multiple cloning sites)
.")
17
L1 The lac operon — cAMP receptor protein cAMP receptor protein cAMP receptor protein(CRP)is a transcriptional activator which is activated by binding to cAMP. However, it is only active when cAMP bound, and cAMP is controlled by glucose. CRP activator mediates the global regulation of gene expression from catabolic operons in response to glucose levels.

18
The P lac is a weak promoter, lacking a strong – 35 and –10 consensus sequences. High level expression from this promoter requires the activity of the specific activator, CRP.

19
When glucose is present The level of cAMP is low in cell, and CRP exists as a dimer which can’t bind to DNA to regulate transcription. When glucose is absent The level of cAMP increase and CRP bind to cAMP. The CRP-cAMP complex binds to P lac just upstream from the site for RNA polymerase. Induces a 90°bend in DNA which enhances RNA polymerase binding to the promoter and thus the transcription by 50-fold.

20
CRP-binding site is an inverted repeat.

21
C A B Summary A: RNA polymerase B: lac repressor C: CRP-cAMP

22
The CRP (also called CAP) protein can bind at different sites relative to RNA polymerase. Supp.
 protein can bind at different sites relative to RNA polymerase. Supp.")
23
L2 The trp operon — The tryptophan operon 1. The trp operon encodes five structural genes required for tryptophan synthesis. 2. It encodes a signal transcription ( 7kb, polycistron ) downstream of Otrp. 3. These genes are co-ordinately expressed when tryptophan is in short supply in the cell.
 downstream of Otrp. 3. These genes are co-ordinately expressed when tryptophan is in short supply in the cell..")
25
L2 The trp operon — The trp repressor 1. Trp repressor is encoded by a separate operon trpR, and specifically interacts with O trp, a palindrome of 18 bp, and overlaps with the P trp sequence between base –21 and +3)
.")
26
2. The repressor can only bind to the operator O trp when it is complexed with tryptophan. Therefore, try is a co- repressor and inhibits its own synthesis through end-product inhibition (negative feed-back regulation).
..")
27
3. The repressor reduces transcription initiation by around 70-fold, which is much smaller than the binding of lac repressor. 4. The repressor is a dimer of two subunits which has a structure with a central core and two flexible DNA-reading heads (carboxyl-terminal of each subunit )
.")
28
trpR operon trp operon

29
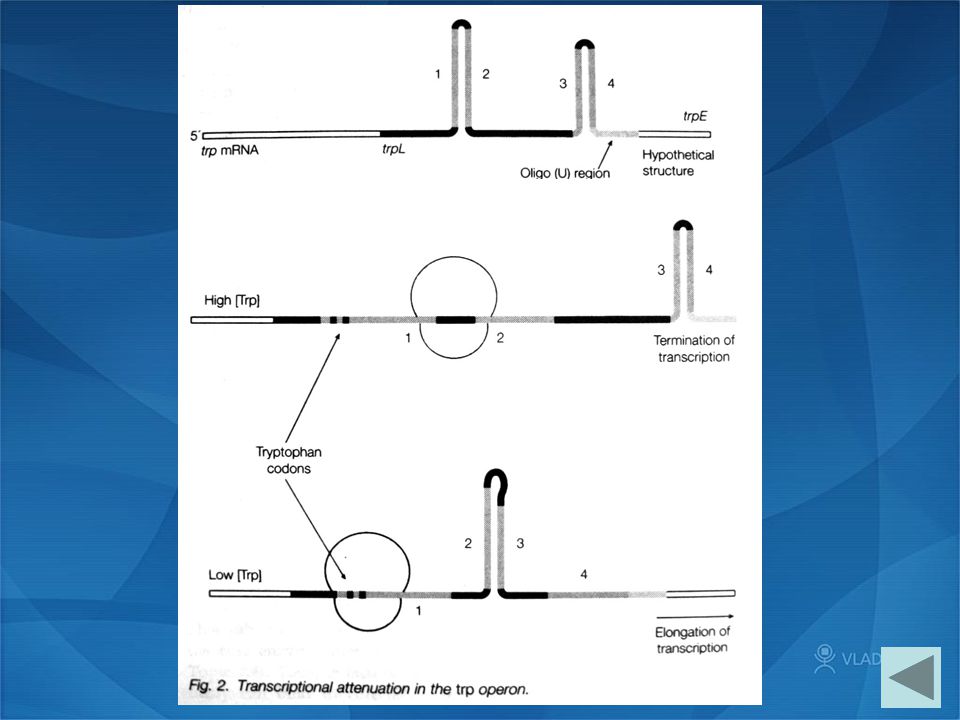
L2 The trp operon — The attenuator Complementary 3:4 termination of transcription Complementary 2:3 Elongation of transcription free leader RNA

30
L2 The trp operon — Leader RNA structure The trp leader RNA contains four regions of complementary sequence which are capable of forming alternative hairpin structure. One of these structures is the attenuator hairpin.

31
L2 The trp operon — The leader peptide The leader RNA contains an efficient ribosome binding site (RBS) and encodes a 14-amino-acid leader peptide (bases 27- 68), Codons 10 and 11 of this peptide encode trp. Thus the availability of trp will affect the translation/ ribosome position, which in turn to regulate transcription termination.

34
L2 The trp operon — Attenuation Transcription and translation in bacteria are coupled. Therefore, synthesis of the leader peptide immediately follows the transcription of leader RNA, and the attenuation is possible

35
High trp Trp is inserted at the trp codons Translate to the end of leader message Ribosome occlude sequence 2 Terminate transcription because 3:4 hairpin formed

36
Lack of trp Lack of aminoacyl tRNA phe Ribosome pause at trp codons, occluding sequence 1 2:3 hairpin (anti-terminator ) forms Transcription into trpE and beyond
 forms Transcription into trpE and beyond")
37
L2 The trp operon — Importance of attenuation A typical negative feed-back regulation Give rise to a 10-fold repression of the trp operon transcription, increasing the regulatory effect up to 700-fold combining the 70-fold repressor effect. Faster and more subtle regulation of trp metabolism in bacteria.

38
L3 Transcriptional regulation by alternative σ factor — Sigma factor σ factor subunit bound to RNA pol for transcription initiation Released core enzyme αββ’ω RNA elongation σfactors is bifunctional protein Bind to core RNA Pol Recognize specific promoter sequence (-35 and –10) in DNA
 in DNA")
40
L3 Transcriptional regulation by alternative σ factor — Promoter recognition In E.coli, σ 70 is responsible for recognition of the -10 and -35 consensus sequence. Differing consensus sequence are found in sets of genes which are regulated by the use of alternative σ factors.

41
L3 Transcriptional regulation by alternative σ factor — Heat shock The response to heat shock is one example in E. coli where gene expression is altered significantly by the use of different s factors.

42
Heat shock Transiently expression of the 17 heat shock proteins Increase in temperature is more extremely (50ºC) Heat shock proteins are the only proteins made in E. coli to maintain its viability From 37ºC to 42ºC

43
L3 Transcriptional regulation by alternative σ factor — Sporulation in Bacteriophage subtilis Under non-optimal environmental conditions Bacillus subtilis cells from spores through a basic cell differentiation process involving cell partitioning into mother cell and forespore.

45
L3 Transcriptional regulation by alternative σ factor — Baxteriophage σ factors Many bacteriophages synthesize their own factors in order to ‘take over’ the host cell’s transcription machinery by substituting the normal cellular factor and altering the promoter specificity of the RNA polymerase.

46
Multiple choice questions 1. Which two of the following statements are correct? A the double stranded DNA sequence that has the upper strand sequence 5'- GGATCGATCC-3' is a palindrome. B the double stranded DNA sequence that has the upper strand sequence 5'- GGATCCTAGG-3' is apalindrome. C the Lac repressor inhibits binding of the polymerase to the lac promoter. D the lac operon is directly induced by lactose. E binding of Lac repressor to allolactose reduces its affinity for the lac operator. F IPTG is a natural inducer of the lac promoter. 2. Which one of the following statements about catabolite-regulated operons is false? A cAMP receptor protein (CRP) and catabolite activator protein (CAP) are different names for the same protein. B when glucose is present in the cell cAMP levels fall. C CRP binds to cAMP and as a result activates transcription. D CRP binds to DNA in the absence of cAMP. E CRP can bend DNA, resulting in activation of transcription.
 and catabolite activator protein (CAP) are different names for the same protein. B when glucose is present in the cell cAMP levels fall. C CRP binds to cAMP and as a result activates transcription. D CRP binds to DNA in the absence of cAMP. E CRP can bend DNA, resulting in activation of transcription..")
47
3. Which one of the following statements about the trp operon is true? A the RNA product of the trp operon is very stable. B the Trp repressor is a product of the trp operon. C the Trp repressor , like the Lac repressor, is a tetramer of identical subunits. D the Trp repressor binds to tryptophan. E tryptophan activates expression from the trp operon. F the trp operon is only regulated by the Trp represso 4. Which two of the following statements about attenuation at the trp operon are true? A attenuation is rho-dependent. B deletion of the attenuator sequence results in an increase in both basal and activated levels of tran- scription from th~ trp promoter. C the attenuator lies upstream of the trp operator sequence. D attenuation does not require tight coupling between transcription and translation. E pausing of a ribosome at two tryptophan codons in the leader peptide when tryptophan is in short supply causes attenuation. F a hairpin structure called the pnti-terminator stops formation of the terminator hairpin, resulting in transcriptional read-through into the trpE gene, when tryptophan is scarce.

48
5. Which two of the following statements about sigma factors are false? A the E. coli RNA polymerase core enzyme cannot start transcription from promoters in the absence of a sigma factor subunit. B different sigma factors may recognize different sets of promoters. C sigma factors recognize both the -10 and -35 promoter elements. D heat shock promoters in E. coli have different -35 and -10 sequences and bind to a diverse set of 17 heat shock sigma factors. E sporulation in B. subtilis is regulated by a diverse set of sigma factors. F bacteriophage T7 expresses its own set of sigma factors as an alternative to encoding its own RNA polymerase.

49
THANK YOU !

Similar presentations











 Do each of his cells have the same genes? Yes, with an exception: germ cells are haploid.>")













