Download presentation
Presentation is loading. Please wait.

1
BIOL 4120: Principles of Ecology Lecture 10: Temporal And Spatial Dynamics of Populations Dafeng Hui Office: Harned Hall 320 Phone: 963-5777 Email: dhui@tnstate.edu

2
Temporal and Spatial Dynamics of populations Numbers of gyrfalcons exported from Iceland to Denmark during 1731 and 1770 reflected population cycles.

3
Topics (Chapter 12) 10. 1 Fluctuation is the rule for natural populations 10.2 Temporal variation affects the age structure of populations 10.3 Population cycles result from time delays in the response of populations to their own density 10.4 Metapopulations are discrete subpopulations linked by movements of individuals 10.5 Chance event may cause small population to go extinct

4
10.1 Fluctuation is the rule for natural populations Domestic sheep on the island of Tasmania of Australia. Introduced in early 1800s and reached the K in about 30 years and varied slightly

6
What are the causes? Sensitivity to environmental change and response time of the population Sheep: large, greater capacity for homeostasis, and better resist physical changes long life, generation overlap, even out the short-term fluctuation in birth rate long life, generation overlap, even out the short-term fluctuation in birth rate Algae and diatom: short life span (a few days), rapid turn over, high mortality, population size depends on continued reproduction, which is sensitivity to food availability, predation, and physical conditions. Phytoplankton populations are intrinsically unstable.
, rapid turn over, high mortality, population size depends on continued reproduction, which is sensitivity to food availability, predation, and physical conditions. Phytoplankton populations are intrinsically unstable..")
7
Periodic cycles of some species Population cycles of grouse in Finland are synchronized across species and areas (Three species in two areas). (6-7 year cycles)
.")
8
Populations of four moth species in the same habitat fluctuate independently

9
10.2 Temporal variation affects the age structure of populations Variation in population size over time often leaves its mark on age structure Variation in population size over time often leaves its mark on age structure Age structure influences the rate of population growth Age structure influences the rate of population growth Commercial whitefish in Lake Erie Commercial whitefish in Lake Erie 1944 resulted a large population 1944 resulted a large population

10
Age distributions of forest trees show the effects of disturbances on seedling establishment (tree ring count) Survey in Pennsylvania, 1928 over the past 400 years
 Survey in Pennsylvania, 1928 over the past 400 years")
11
10.3 Population cycles result from time delays in the response of populations to their own density Cycling of populations has been observed Cycling of populations has been observed E.g: hare cycles, 11-yr cycle in sunspot E.g: hare cycles, 11-yr cycle in sunspot Oscillation and time delays Oscillation and time delays Oscillation may reflect intrinsic dynamic qualities of biological systems (some oscillate even with small environmental fluctuation) Oscillation may reflect intrinsic dynamic qualities of biological systems (some oscillate even with small environmental fluctuation) Time delays in responses: high birth rate -> overshoot population -> high death rate -> low population. Time delays in responses: high birth rate -> overshoot population -> high death rate -> low population.

12
Time delays and oscillations in discrete- time models ΔN(t)=N(t+1)-N(t) =RN(t) R is proportional increase or decrease in N per unit of time Let’s make R density- dependent, ΔN(t)=RN(t) (1-N(t)/K) =RN(t)/K (K-N(t)) K-N(t) is the difference between the size of the population and its carrying capacity at time t Damped oscillation, limit cycle or chaos
=N(t+1)-N(t) =RN(t) R is proportional increase or decrease in N per unit of time Let’s make R density- dependent, ΔN(t)=RN(t) (1-N(t)/K) =RN(t)/K (K-N(t)) K-N(t) is the difference between the size of the population and its carrying capacity at time t Damped oscillation, limit cycle or chaos")
13
Time delays and oscillations in continuous-time models Logistic model: dN/dt=r N(1-N/K) or dN(t)/dt=rN(t)(1-N(t)/K) (1-N(t)/K) is a component that show density dependent influence by population size If not by current, but by a population size at time (t-τ) dN(t)/dt=rN(t)(1-N(t-τ)/K) Oscillation depends on (rτ): r τ=0, no oscillation r τ=1, damped oscillation, r τ=2, limit cycles
 or dN(t)/dt=rN(t)(1-N(t)/K) (1-N(t)/K) is a component that show density dependent influence by population size If not by current, but by a population size at time (t-τ) dN(t)/dt=rN(t)(1-N(t-τ)/K) Oscillation depends on (rτ): r τ=0, no oscillation r τ=1, damped oscillation, r τ=2, limit cycles")
14
Cycles in laboratory populations Water flea experiment 25oC, large oscillation period of cycle: 60 days time delays: 12-15 days (average age give birth: 12-15 days at 25oC) 18oC, no oscillation reproduction fell off quickly with increasing density, and life span was longer than at 25oC. Deaths were more evenly distributed over all ages and some individuals gave birth even at high population densities. Generation overlap broadly, No time delay.

15
Introduction of time delays results in regular population cycles Blowfly experiment AJ Nicholson, Australian Control population by provide limited food supplies to larvae and unlimited food to adults. Adults population cycles from 0 to 4000. Period 30-40 days Cause: a time delay in the responses of fecundity and mortality to the density of adults. A time delayed logistic model (rt=2.1) provides a good fit for the blowfly data.
 provides a good fit for the blowfly data..")
16
Limited food supplies to adults limited the time delay and results in the elimination of population cycles What happens if you do not provide enough food for adults?

17
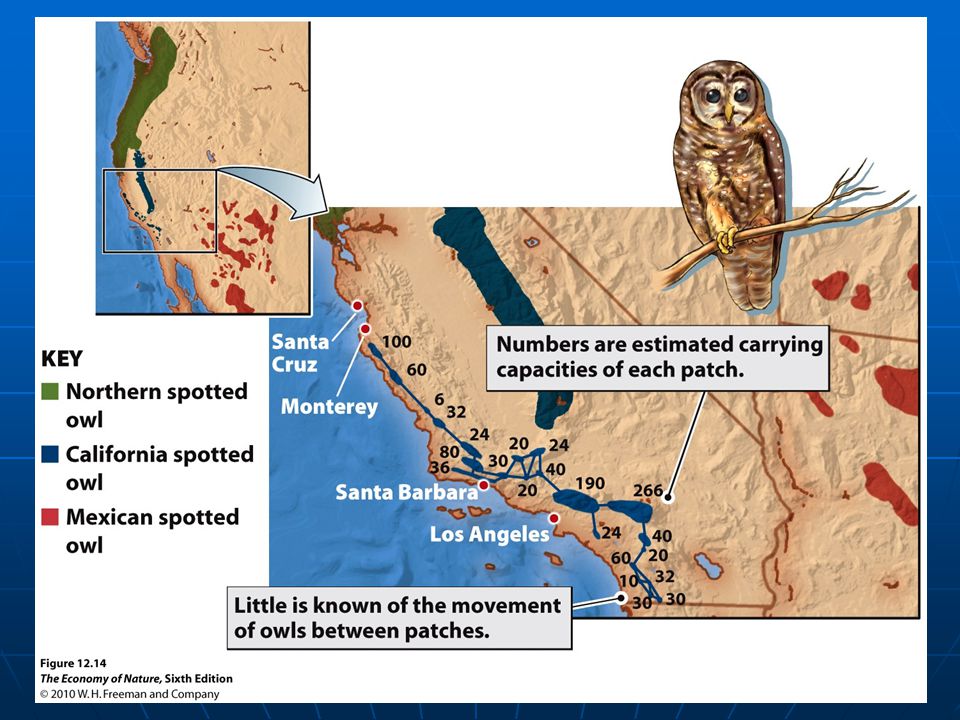
10.4 Metapopulations are discrete subpopulation linked by movements of individuals Habitat Patches, subpopulations and metapopulation Habitat Patches, subpopulations and metapopulation Processes contribute to dynamics of metapopulations Processes contribute to dynamics of metapopulations Growth and regulation of subpopulations within patchesGrowth and regulation of subpopulations within patches Colonization of empty patches by migrating individuals to form new subpopulationsColonization of empty patches by migrating individuals to form new subpopulations and the extinction of established subpopulations.and the extinction of established subpopulations.

19
Basic model of metapopulation dynamics One population is divided into discrete subpopulations, each subpopulation has a probability of going extinct (e). If (p) is the fraction of suitable habitat patches occupied by subpopulations, then subpopulations go extinct at the rate (ep). The rate of colonization of empty patches depends on the fraction of patches that are empty (1-p) and the fraction of patches sending out potential colonists (p). The rate of colonization within the metapopulation as a whole as a single rate constant (c) times the product (p(1-p)). The rate of change in patch occupancy: dp/dt = cp(1-p) – ep colonization extinction
 is the fraction of suitable habitat patches occupied by subpopulations, then subpopulations go extinct at the rate (ep). The rate of colonization of empty patches depends on the fraction of patches that are empty (1-p) and the fraction of patches sending out potential colonists (p). The rate of colonization within the metapopulation as a whole as a single rate constant (c) times the product (p(1-p)). The rate of change in patch occupancy: dp/dt = cp(1-p) – ep colonization extinction.")
20
Basic model of metapopulation dynamics The rate of change in patch occupancy: dp/dt = cp(1-p) – ep colonization extinction A metapopulation attains equilibrium size when colonization equals extinction cp(1-p) = ep Thus, p^= 1 – e/c This is the proportion of occupied patches at metapopulation equilibrium
 – ep colonization extinction A metapopulation attains equilibrium size when colonization equals extinction cp(1-p) = ep Thus, p^= 1 – e/c This is the proportion of occupied patches at metapopulation equilibrium")
21
Recap Population cycles: very common Mechanisms: result from time delays in the response of populations to their own density Two models: how R or rt influences the population size change. Two models: how R or rt influences the population size change. Metapopulations and subpopulations Basic model of metapopulation dynamics Basic model of metapopulation dynamics

22
Basic model of metapopulation dynamics dp/dt = cp(1-p) – ep colonization extinction p^= 1 – e/c Rate of e/c is very important If e = 0, then p^=1, all patches occupied, none disappears If e>=c, then p^ =0, then metapopulation heads toward extinction 0< e < c, intermediate value, results in a shifting mosaic of occupied and unoccupied patches.
 – ep colonization extinction p^= 1 – e/c Rate of e/c is very important If e = 0, then p^=1, all patches occupied, none disappears If e>=c, then p^ =0, then metapopulation heads toward extinction 0< e < c, intermediate value, results in a shifting mosaic of occupied and unoccupied patches.")
23
Basic model of metapopulation dynamics In this model, there are many assumptions 1.All patches are equal 2.(e) and (c) for each patch are the same 3.Each occupied patch contribute equally to dispersal 4.Colonization and extinction in each patch occur independently of other patches 5.Colonization rate is proportional to the fraction of occupied patches (p) In reality: Patches vary in size, habitat quality, degree of isolation from other patches. Larger patch support large subpopulation, lower probability of extinction. Smaller, more isolated patches are less likely to be occupied.

24
Larger, less isolated patches are more likely to be occupied Shrew on islands in two lakes in Finland Few occupied patch area <1 Isolation?

25
Larger, less isolated patches are more likely to be occupied Butterfly on patches of calcareous grassland in England (Hanski et al. 1991) Patch size and isolation are important
 Patch size and isolation are important.")
26
Glanville fritillary butterfly study by Illka Hanski, Finland A survey for p^: Occupied patches of dry meadows, Aland Islands, Finland Exp. One: Total 1600 suitable patches, only 30% were occupied at any given time. Exp. Two: Introduced populations to 10 of the 20 suitable habitat patches on the smaller, isolated island of Scottungia, in August 1991 Observed over next 10 years Number of extinctions varies between 0 to 12 per year Number of colonization between 0 and 9 Subpopulations: started at 10, dropped to as few as 2, and increased as high as 14, ended at 11. None of the original 10 survived the decade, the metapopulation as a whole persisted.

27
The rescue effect Immigration from large, productive subpopulations can keep declining subpopulations (small ones) from dwindling to small numbers and eventually becoming extinct. This phenomenon is known as rescue effect. Dispersal is critical for colonizing empty patches, as well as maintaining established populations. Model: modify the model and add rate of extinction (e) decrease as the fraction of occupied patches (p) increases (more rescuers) dP/dt = cp(1-p) - ep(1-p) p=0, 1 if c><e. This model predicts that p^ will either equal to 0, otherwise, it will increase to 1, as when p<1 but close to 1, 1-p is small, reduce the extinction rate. With rescue effect, all patches will be occupied.
 decrease as the fraction of occupied patches (p) increases (more rescuers) dP/dt = cp(1-p) - ep(1-p) p=0, 1 if c><e. This model predicts that p^ will either equal to 0, otherwise, it will increase to 1, as when p<1 but close to 1, 1-p is small, reduce the extinction rate. With rescue effect, all patches will be occupied..")
28
10.5 Chance event may cause small populations to go extinction Deterministic model and stochastic model: Population models we described before are based on average values of birth rate and death rate, and assume no difference among individuals. Such models, whose outcomes can be predicted with certainty, are called deterministic models. Models built in with chance factors (randomness), such as birth and death rate vary from each individual to another and from one time to another (with mean over certain time or of all individuals is fixed). The result of each model run varies and can’t be predicted, stochastic model.
, such as birth and death rate vary from each individual to another and from one time to another (with mean over certain time or of all individuals is fixed). The result of each model run varies and can’t be predicted, stochastic model..")
29
Three types of randomness 1. Unpredictable catastrophe, such as appearance of a predator, disease, fire etc (birth and death) 2. Environmental variation (some rules, small variation not predictable). Physical and environmental factors (influence birth and death) 3. Stochastic processes such as death of an individual, number of offspring produced by an individual. Even under constant environment, these values could change for an individual. (Overall, there is a probability distribution) (coin tossing is an example of a stochastic process)
 2. Environmental variation (some rules, small variation not predictable). Physical and environmental factors (influence birth and death) 3. Stochastic processes such as death of an individual, number of offspring produced by an individual. Even under constant environment, these values could change for an individual. (Overall, there is a probability distribution) (coin tossing is an example of a stochastic process).")
30
Stochastic population processes produce a probability distribution of population size N(0)=10, lambda=1.5 No stochastic process involved, what’s the population size at t=1? Chart on left: N=10, pure birth, b=0.5 and stochastic process involved, All give birth, N1=10+10= 20.

31
Chance events exert their influence more forcefully in small population than in large ones Coin tossing: a set of 5 pennies, 5 heads in a trial is 1/32; 10 pennies, is 1/1024. If each individual in a population is a coin, and heads mean death, a population of 5 has a higher probability of extinction, just by chance.

32
Stochastic extinction of small populations Random walk: A population subject to stochastic birth and death process is said to take a random walk, meaning that its numbers may increase or decrease strictly by chance. When the size of such a population does not respond to changes in density, its ultimate fate is extinction, regardless of how its size might increase in the meantime. Mathematicians have calculated the probability of extinction. For simplicity, given same birth rate and death rate, the probability of a population will die out within a time interval t is p(t)=[bt/(1+bt)]^N.
=[bt/(1+bt)]^N..")
33
Stochastic extinction of small populations The probability of stochastic extinction increase over time, but decrease as a function of initial population size N. b=0.5, p0(t)=[bt/(1+bt)] ^N N=10, t=10, p=0.16 t=1,000, p=0.98
=[bt/(1+bt)] ^N N=10, t=10, p=0.16 t=1,000, p=0.98.")
34
Stochastic extinction with density dependence Stochastic extinction models usually do not include density- dependent changes in birth and death rates. If density-dependent birth and death rate includes, it rarely goes extinction (unless the population size is very small), as a population drops below K, the birth rate will increase and death rate will decrease. Whether the density-independent stochastic models are relevant to natural populations? They are. 1)Fragmentation by human beings creates many small subpopulation, often so isolated that eventual demise can’t prevented by immigration from other populations 2)Changing environmental conditions reduce fecundity 3)Endangered species can’t compete with other species 4)Small populations sometimes exhibit positive density dependence (Allee effect), their number may decline more rapidly.
, as a population drops below K, the birth rate will increase and death rate will decrease. Whether the density-independent stochastic models are relevant to natural populations. They are. 1)Fragmentation by human beings creates many small subpopulation, often so isolated that eventual demise can’t prevented by immigration from other populations 2)Changing environmental conditions reduce fecundity 3)Endangered species can’t compete with other species 4)Small populations sometimes exhibit positive density dependence (Allee effect), their number may decline more rapidly..")
35
Size and extinction of natural populations Small size populations become more susceptible to extinction, particularly on small islands An Example: species lists for birds in 1917 and 1968 on two islands. Over 51 years, 10 species disappeared from Santa Barbara Island (3 km^2), only 6 of 36 disappeared from the large Santa Cruz Island (249 km^2) Extinction rate: 1.7% and 0.1% per year.
, only 6 of 36 disappeared from the large Santa Cruz Island (249 km^2) Extinction rate: 1.7% and 0.1% per year..")
36
The End

38
10.9 Population extinction If r becomes negative (birth rate < death rate), population declines and will go extinction. If r becomes negative (birth rate < death rate), population declines and will go extinction. Factors: Extreme environmental events (droughts, floods, cold or heat etc), loss of habitat (human). Factors: Extreme environmental events (droughts, floods, cold or heat etc), loss of habitat (human). Small populations are susceptible to extinction Small populations are susceptible to extinction Allee effect, genetic drift, inbreeding (mating between relatives) Overgraze, only 8 in 1950
, population declines and will go extinction. Factors: Extreme environmental events (droughts, floods, cold or heat etc), loss of habitat (human). Factors: Extreme environmental events (droughts, floods, cold or heat etc), loss of habitat (human). Small populations are susceptible to extinction Small populations are susceptible to extinction Allee effect, genetic drift, inbreeding (mating between relatives) Overgraze, only 8 in")
39
Small population size may result in the breakdown of social structures that are integral to successful cooperative behaviors (mating, foraging, defense) Small population size may result in the breakdown of social structures that are integral to successful cooperative behaviors (mating, foraging, defense) The Allee effect is the decline in reproduction or survival under conditions of low population density The Allee effect is the decline in reproduction or survival under conditions of low population density There is less genetic variation in a small population and this may affect the population’s ability to adapt to environmental change There is less genetic variation in a small population and this may affect the population’s ability to adapt to environmental change
 Small population size may result in the breakdown of social structures that are integral to successful cooperative behaviors (mating, foraging, defense) The Allee effect is the decline in reproduction or survival under conditions of low population density The Allee effect is the decline in reproduction or survival under conditions of low population density There is less genetic variation in a small population and this may affect the population’s ability to adapt to environmental change There is less genetic variation in a small population and this may affect the population’s ability to adapt to environmental change")
40
Hackney and McGraw (West Virginia University) examined the reproductive limitations by small population size on American ginseng (Panax quinquefolius) Hackney and McGraw (West Virginia University) examined the reproductive limitations by small population size on American ginseng (Panax quinquefolius) Fruit production per plant declined with decreasing population size due to reduced visitation by pollinationFruit production per plant declined with decreasing population size due to reduced visitation by pollination
 examined the reproductive limitations by small population size on American ginseng (Panax quinquefolius) Hackney and McGraw (West Virginia University) examined the reproductive limitations by small population size on American ginseng (Panax quinquefolius) Fruit production per plant declined with decreasing population size due to reduced visitation by pollinationFruit production per plant declined with decreasing population size due to reduced visitation by pollination")
42
Recap Population cycles: very common Mechanisms: result from time delays in the response of populations to their own density Two models: how R or rt influences the population size change. Two models: how R or rt influences the population size change. Metapopulations and subpopulations Basic model of metapopulation dynamics Basic model of metapopulation dynamics

Similar presentations



 (red, orange, green) U.>")


 & deaths (D)>")






 How many are in the population? 2) Are its numbers increasing.>")







