Download presentation
Presentation is loading. Please wait.

1
Principles of Cell Communication

2
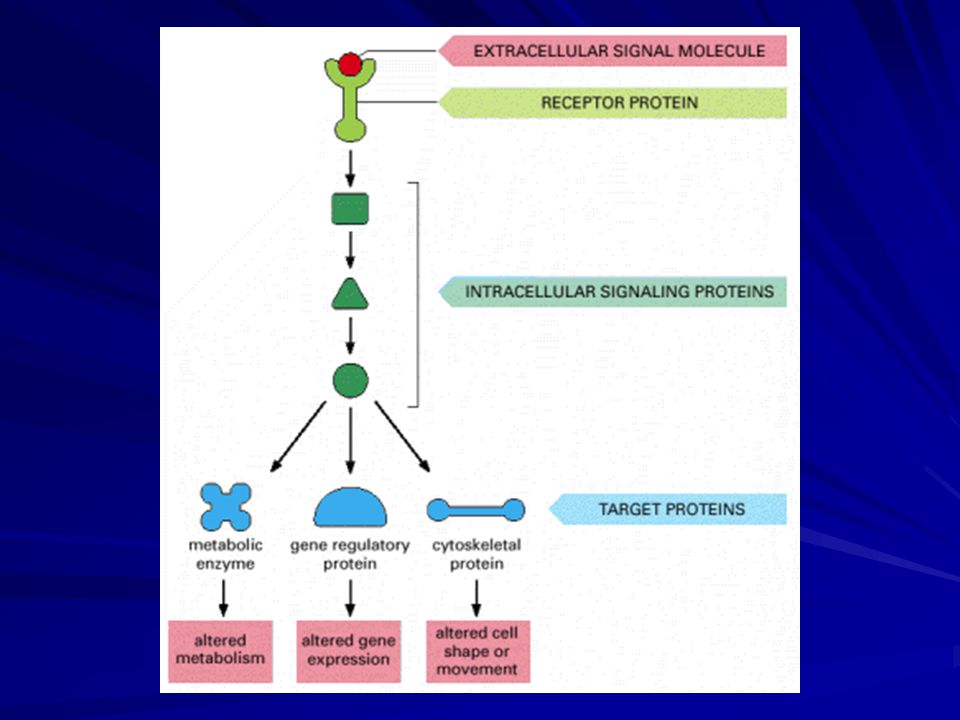
Extracellular signal molecules bind to specific receptors
Cells communicate by hundreds of signal molecules. These include proteins, small peptides, amino acids, nucleotides, steroids, retinoids, fatty acids, and dissolved gases. The target cell responds by means of a specific protein called a receptor. It binds the signal molecule and then initiates a response.

3
The extracellular signal molecule often act at very low concentrations (<10^-8 M) and the receptors that bind them have a high affinity (Ka > 10^8 L/M). These receptors are transmembrane proteins on the target cell surface. When they bind an extracellular signal molecule (a ligand), they become activated and generate a cascade of intracellular signals that alter the behaviour of the cell.
, they become activated and generate a cascade of intracellular signals that alter the behaviour of the cell.")
5
Different cells can respond differently to the same extracellular signal molecule
The specific way in which a cell reacts to its environment varies. It varies according to the set of receptor proteins the cell possesses, which determines the particular subset of signals it can respond to, and it varies according to the intracellular machinery by which the cell integrates and interprets the signals it receives.

6
Thus, a single signal molecule often has different effects on different target cells.
The same signal molecule bind to identical receptor proteins yet produces very different responses in different types of target cells, reflecting differences in the internal machinery to which the receptors are coupled.

8
Nitric oxide signals by binding directly to an enzyme inside the target cell
Although most extracellular signals are hydrophilic molecules that bind to receptors on the surface of the target cell, some signal molecules are hydrophobic enough and small enough to pass readily across the target cell-plasma membrane. NO functions to regulate smooth muscle contractions, for nerve cells to signal to their neighbors, and is produced as a local mediator by macrophages and neutrophils to assist in killing microbes. NO is made by the deamination of arginine, catalyzed by NO synthase.

9
Because it passes across membranes, NO rapidly diffuses out of the cell and into neighboring cells.
It acts locally because it has a half-life of 5-10 seconds (converted to nitrates and nitrites). NO binds to iron in the active site of the enzyme guanylyl cyclase to produce cGMP inducing vasodilatation. CO is another gas that is used as an intercellular signal. It can act in the same way as NO, by stimulating guanylyl cyclase.
. NO binds to iron in the active site of the enzyme guanylyl cyclase to produce cGMP inducing vasodilatation. CO is another gas that is used as an intercellular signal. It can act in the same way as NO, by stimulating guanylyl cyclase.")
11
Nuclear receptors are ligand-activated gene regulatory proteins
A number of small hydrophobic signal molecules diffuse directly across the plasma membrane and bind to intracellular receptor proteins. These signal molecules include steroid hormones, thyroid hormones, retinoids, and vitamin D. When these signals bind to their receptors they bind to DNA to regulate the transcription of specific genes. These signal molecules are insoluble and are made soluble for transport in the bloodstream by binding to carrier proteins.

12
Steroid hormones persist in the blood for hours.
Therefore, water-soluble signal molecules usually mediate responses of short duration whereas insoluble ones tend to mediate responses that are longer lasting. The intracellular receptors for these molecules all bind to specific DNA sequences adjacent to the genes that the ligand regulates. Some receptors are located in the cytosol and enter the nucleus after binding; others are bound directly to DNA without a ligand.

13
The ligand binding also causes the receptor to bind to coactivator proteins that induce gene transcription. The transcription process response usually takes place in successive steps: the direct activation of a small number of specific genes occurs within about 30 minutes and constitutes the primary response; the protein products of these genes in turn activate other genes to produce a delayed, secondary response; and so on. The responses to steroid and thyroid hormones, vitamin D, and retinoids, like responses to extracellular signals in general, are determined as much by the nature of the target cell as by the nature of the signal molecule.

14
The Three Largest Classes of Cell-Surface Receptor Proteins Are Ion-Channel-linked, G-Protein-linked, and Enzyme-linked Receptors All water-soluble signal molecules (including neurotransmitters and all signal proteins) bind to specific receptor proteins on the surface of the target cells that they influence. These cell-surface receptor proteins act as signal transducers.
 bind to specific receptor proteins on the surface of the target cells that they influence. These cell-surface receptor proteins act as signal transducers.")
15
Most cell-surface receptor proteins belong to one of three classes.
Ion-channel-linked receptors, also known as transmitter-gated ion channels or ionotropic receptors, are involved in rapid synaptic signaling between electrically excitable cells. G-protein-linked receptors act indirectly to regulate the activity of a separate plasma-membrane-bound target protein, which can be either an enzyme or an ion channel. Enzyme-linked receptors, when activated, either function directly as enzymes or are directly associated with enzymes that they activate.

17
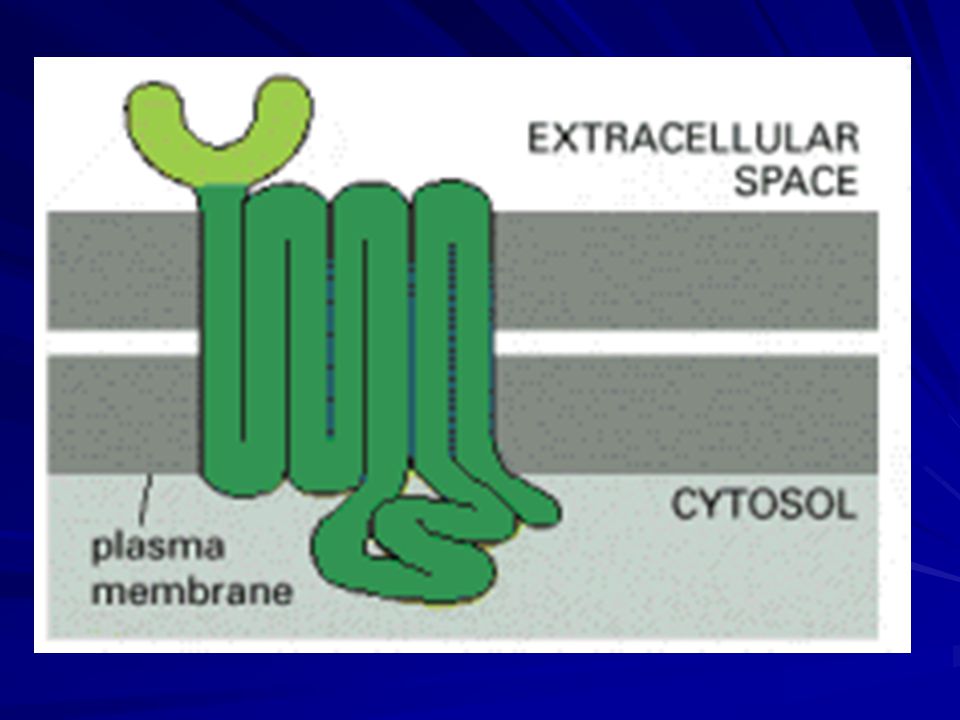
Signaling through G-Protein-Linked Cell-Surface Receptors
G-protein-linked receptors form the largest family of cell-surface receptors and are found in all eukaryotes. Despite the chemical and functional diversity of the signal molecules that bind to them, all G-protein-linked receptors have a similar structure. They consist of a single polypeptide chain that threads back and forth across the lipid bilayer seven times and are therefore sometimes called serpentine receptors.

19
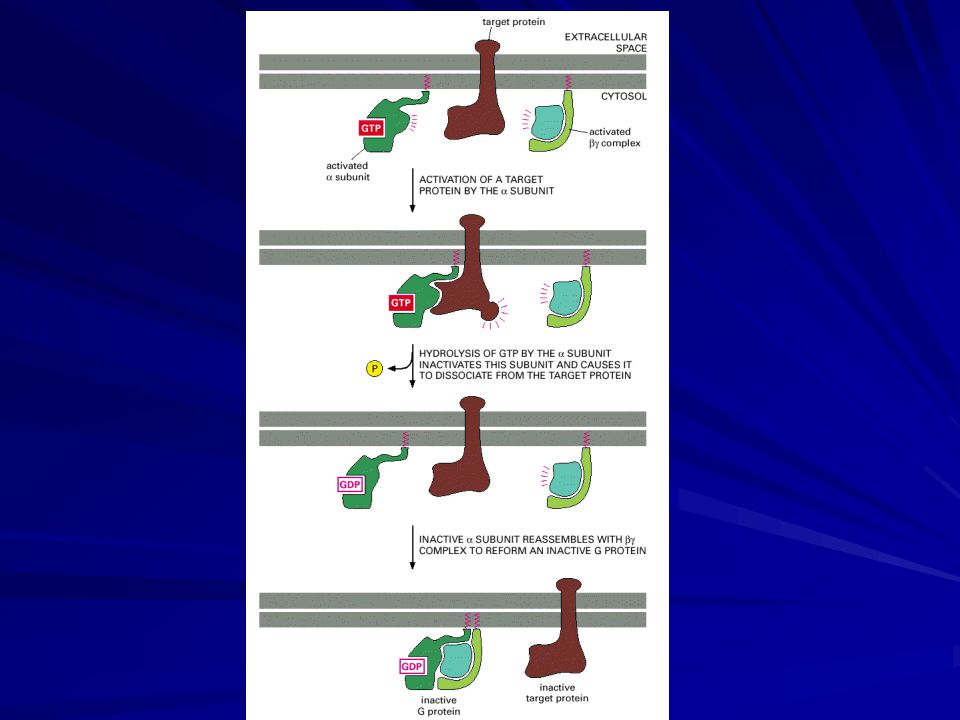
Trimeric G Proteins Disassemble to Relay Signals from G-Protein-linked Receptors
When extracellular signaling molecules bind to serpentine receptors, the receptors undergo a conformational change that enables them to activate trimeric GTP-binding proteins (G proteins). These G proteins are attached to the cytoplasmic face of the plasma membrane, where they serve as relay molecules, functionally coupling the receptors to enzymes or ion channels in this membrane. G proteins are composed of three protein subunits—α, β, and γ. In the unstimulated state, the α subunit has GDP bound and the G protein is inactive. When stimulated by an activated receptor, the α subunit releases its bound GDP, allowing GTP to bind in its place.
. These G proteins are attached to the cytoplasmic face of the plasma membrane, where they serve as relay molecules, functionally coupling the receptors to enzymes or ion channels in this membrane. G proteins are composed of three protein subunits—α, β, and γ. In the unstimulated state, the α subunit has GDP bound and the G protein is inactive. When stimulated by an activated receptor, the α subunit releases its bound GDP, allowing GTP to bind in its place.")
21
This exchange causes the trimer to dissociate into two activated components—an α subunit and a βγ complex. GTP binding causes a conformational change that affects the surface of the α subunit that associates with the βγ complex in the trimer. This change causes the release of the βγ complex, but it also causes and the α subunit to adopt a new shape that allows it to interact with its target proteins. The βγ complex does not change its conformation, but the surface previously masked by the α subunit is now available to interact with a second set of target proteins.

23
The α subunit is a GTPase, and once it hydrolyzes its bound GTP to GDP, it reassociates with a βγ complex to re-form an inactive G protein, reversing the activation process. The time during which the α subunit and βγ complex remain apart and active is usually short, and it depends on how quickly the α subunit hydrolyzes its bound GTP. The GTPase activity of the α subunit is greatly enhanced by the binding of a second protein, which can be either its target protein or a specific modulator known as a regulator of G protein signaling (RGS).
.")
24
Some G Proteins Signal By Regulating the Production of Cyclic AMP
Cyclic AMP is synthesized from ATP by a plasma-membrane-bound enzyme adenylyl cyclase, and it is rapidly and continuously destroyed by one or more cyclic AMP phosphodiesterases that hydrolyze cyclic AMP to adenosine 5′-monophosphate (5′-AMP). Many extracellular signal molecules work by increasing cyclic AMP content, and they do so by increasing the activity of adenylyl cyclase rather than decreasing the activity of phosphodiesterase.
. Many extracellular signal molecules work by increasing cyclic AMP content, and they do so by increasing the activity of adenylyl cyclase rather than decreasing the activity of phosphodiesterase.")
25
Adenylyl cyclase is a large multipass transmembrane protein with its catalytic domain on the cytosolic side of the plasma membrane. All receptors that act via cyclic AMP are coupled to a stimulatory G protein (Gs), which activates adenylyl cyclase and thereby increases cyclic AMP concentration. Another G protein, called inhibitory G protein (Gi), inhibits adenylyl cyclase, but it mainly acts by directly regulating ion channels rather than by decreasing cyclic AMP content.
, which activates adenylyl cyclase and thereby increases cyclic AMP concentration. Another G protein, called inhibitory G protein (Gi), inhibits adenylyl cyclase, but it mainly acts by directly regulating ion channels rather than by decreasing cyclic AMP content.")
26
Cyclic-AMP-dependent Protein Kinase (PKA) Mediates Most of the Effects of Cyclic AMP
Although cyclic AMP can directly activate certain types of ion channels in the plasma membrane of some highly specialized cells, in most animal cells it exerts its effects mainly by activating cyclic-AMP-dependent protein kinase (PKA). This enzyme catalyzes the transfer of the terminal phosphate group from ATP to specific serines or threonines of selected target proteins, thereby regulating their activity. In the inactive state, PKA consists of a complex of two catalytic subunits and two regulatory subunits.
. This enzyme catalyzes the transfer of the terminal phosphate group from ATP to specific serines or threonines of selected target proteins, thereby regulating their activity. In the inactive state, PKA consists of a complex of two catalytic subunits and two regulatory subunits.")
27
The binding of cyclic AMP to the regulatory subunits alters their conformation, causing them to dissociate from the complex. The released catalytic subunits are thereby activated to phosphorylate specific substrate protein molecules.

28
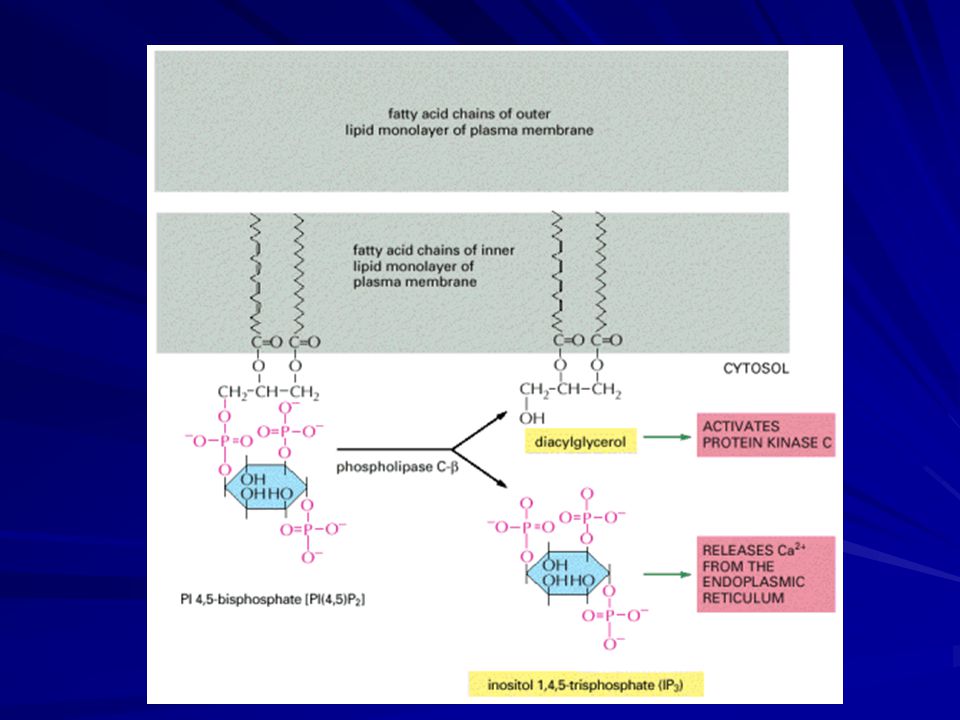
Some G Proteins Activate the Inositol Phospholipid Signaling Pathway by Activating Phospholipase C-β
Many G-protein-linked receptors exert their effects mainly via G proteins that activate the plasma-membrane-bound enzyme phospholipase C-β. The phospholipase acts on an inositol phospholipid (a phosphoinositide) called phosphatidylinositol 4,5-bisphosphate [PI(4,5)P2], which is present in small amounts in the inner half of the plasma membrane lipid bilayer. The activated phospholipase cleaves PI(4,5)P2 to generate two products: inositol 1,4,5-trisphosphate and diacylglycerol. At this step, the signaling pathway splits into two branches.
 called phosphatidylinositol 4,5-bisphosphate [PI(4,5)P2], which is present in small amounts in the inner half of the plasma membrane lipid bilayer. The activated phospholipase cleaves PI(4,5)P2 to generate two products: inositol 1,4,5-trisphosphate and diacylglycerol. At this step, the signaling pathway splits into two branches.")
30
Inositol 1,4,5-trisphosphate (IP3) is a small, water-soluble molecule that leaves the plasma membrane and diffuses rapidly through the cytosol. When it reaches the endoplasmic reticulum (ER), it binds to and opens IP3-gated Ca2+-release channels in the ER membrane. Ca2+ stored in the ER is released through the open channels, quickly raising the concentration of Ca2+ in the cytosol.
, it binds to and opens IP3-gated Ca2+-release channels in the ER membrane. Ca2+ stored in the ER is released through the open channels, quickly raising the concentration of Ca2+ in the cytosol.")
32
Diacylglycerol remains embedded in the membrane, where it has two potential signaling roles.
First, it can be further cleaved to release arachidonic acid, which can either act as a messenger in its own right or be used in the synthesis of other small lipid messengers called eicosanoids. The second, and more important, function of diacylglycerol is to activate a crucial serine/threonine protein kinase called protein kinase C (PKC), so named because it is Ca2+-dependent.
, so named because it is Ca2+-dependent.")
33
The initial rise in cytosolic Ca2+ induced by IP3 alters the PKC so that it translocates from the cytosol to the cytoplasmic face of the plasma membrane. There it is activated by the combination of Ca2+, diacylglycerol, and the negatively charged membrane phospholipid phosphatidylserine. Once activated, PKC phosphorylates target proteins that vary depending on the cell type.

34
Some G Proteins Directly Regulate Ion Channels
In some other cases, G proteins directly activate or inactivate ion channels in the plasma membrane of the target cell, thereby altering the ion permeability—and hence the excitability of the membrane. Acetylcholine released by the vagus nerve, for example, reduces both the rate and strength of heart muscle cell contraction. Once activated, the α subunit of Gi inhibits adenylyl cyclase, while the βγ complex binds to K+ channels in the heart muscle cell plasma membrane to open them.

35
The opening of these K+ channels makes it harder to depolarize the cell, which contributes to the inhibitory effect of acetylcholine on the heart. Other trimeric G proteins regulate the activity of ion channels less directly, either by stimulating channel phosphorylation (by PKA, PKC, or CaM-kinase, for example) or by causing the production or destruction of cyclic nucleotides that directly activate or inactivate ion channels.
 or by causing the production or destruction of cyclic nucleotides that directly activate or inactivate ion channels.")
36
Signaling through Enzyme-Linked Cell-Surface Receptors
Enzyme-linked receptors are a second major type of cell-surface receptor. They were recognized initially through their role in responses to extracellular signal proteins that promote the growth, proliferation, differentiation, or survival of cells in animal tissues. The responses to them are typically slow (on the order of hours) and usually require many intracellular signaling steps that eventually lead to changes in gene expression.
 and usually require many intracellular signaling steps that eventually lead to changes in gene expression.")
37
Enzyme-linked receptors have since been found also to mediate direct, rapid effects on the cytoskeleton, controlling the way a cell moves and changes its shape. Like G-protein-linked receptors, enzyme-linked receptors are transmembrane proteins with their ligand-binding domain on the outer surface of the plasma membrane. Instead of having a cytosolic domain that associates with a trimeric G protein, however, their cytosolic domain either has an intrinsic enzyme activity or associates directly with an enzyme.

38
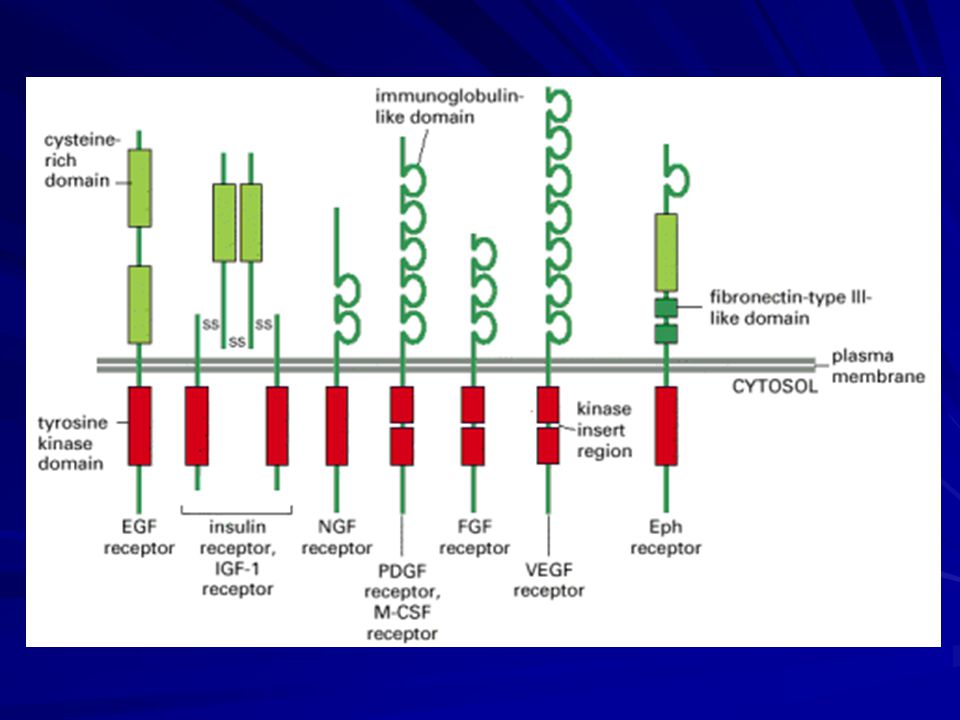
Activated Receptor Tyrosine Kinases Phosphorylate Themselves
The extracellular signal proteins that act through receptor tyrosine kinases consist of a large variety of secreted growth factors and hormones. Notable examples discussed include epidermal growth factor (EGF), platelet-derived growth factor (PDGF), fibroblast growth factors (FGFs), hepatocyte growth factor (HGF), insulin, insulinlike growth factor-1 (IGF-1), vascular endothelial growth factor (VEGF), macrophage-colony-stimulating factor (M-CSF), and all the neurotrophins, including nerve growth factor (NGF). In all cases, the binding of a signal protein to the ligand-binding domain on the outside of the cell activates the intracellular tyrosine kinase domain.
, platelet-derived growth factor (PDGF), fibroblast growth factors (FGFs), hepatocyte growth factor (HGF), insulin, insulinlike growth factor-1 (IGF-1), vascular endothelial growth factor (VEGF), macrophage-colony-stimulating factor (M-CSF), and all the neurotrophins, including nerve growth factor (NGF). In all cases, the binding of a signal protein to the ligand-binding domain on the outside of the cell activates the intracellular tyrosine kinase domain.")
40
Once activated, the kinase domain transfers a phosphate group from ATP to selected tyrosine side chains, both on the receptor proteins themselves and on intracellular signaling proteins that subsequently bind to the phosphorylated receptors. How does the binding of an extracellular ligand activate the kinase domain on the other side of the plasma membrane? For a G-protein-linked receptor, ligand binding is thought to change the relative orientation of several of the transmembrane α helices, thereby shifting the position of the cytoplasmic loops relative to each other. For the enzyme-linked receptors, two or more receptor chains come together in the membrane, forming a dimer or higher oligomer.

41
The rearrangement induced in cytosolic tails of the receptors initiates the intracellular signaling process. For receptor tyrosine kinases, the rearrangement enables the neighboring kinase domains of the receptor chains to cross-phosphorylate each other on multiple tyrosines, a process referred to as autophosphorylation. To activate a receptor tyrosine kinase the ligand usually has to bind simultaneously to two adjacent receptor chains. Autophosphorylation of the cytosolic tail of receptor tyrosine kinases contributes to the activation process in two ways.

42
First, phosphorylation of tyrosines within the kinase domain increases the kinase activity of the enzyme. Second, phosphorylation of tyrosines outside the kinase domain creates high-affinity docking sites for the binding of a number of intracellular signaling proteins in the target cell. Each type of signaling protein binds to a different phosphorylated site on the activated receptor because it contains a specific phosphotyrosine-binding domain that recognizes surrounding features of the polypeptide chain in addition to the phosphotyrosine.

43
Once bound to the activated kinase, the signaling protein may itself become phosphorylated on tyrosines and thereby activated; alternatively, the binding alone may be sufficient to activate the docked signaling protein. In summary, autophosphorylation serves as a switch to trigger the transient assembly of a large intracellular signaling complex, which then broadcasts signals along multiple routes to many destinations in the cell.

Similar presentations










>")














