Download presentation
Presentation is loading. Please wait.

1
Studying adaptation The evolutionary analysis of form and function in natural populations

2
Fundamental questions Is a trait an adaptation? –Not all traits increase fitness (e.g. hemoglobin makes blood red...), although complexity is a key –Chucks by male túngara frogs are not favored by natural selection from D.J. Futuyma, Evolutionary Biology
, although complexity is a key –Chucks by male túngara frogs are not favored by natural selection from D.J. Futuyma, Evolutionary Biology.")
3
Fundamental questions What is the adaptive function (often called adaptive significance) of a trait? –Sometimes this is obvious, other times no –The obvious can be wrong –Finding the truth requires careful evolutionary analysis

4
Example 1:Why are polar bears white? Obvious answer: camouflage when hunting seals Problems with this: –Observations of hunting behavior suggest camouflage is not often used –Most hunts involve ambush at breathing holes (Stirling 1974, Anderson and Guravich 1992)
.")
5
Alternative hypothesis: a solar heat collector Photographed under UV, polar bears are black (they absorb UV) Grojean et al (1980) examined optical properties of polar bear fur –hair is like a “drawn quartz tube with irregular inner surface” –reflects incident UV towards the skin –light scattering and reflection make the fur look white
 Grojean et al (1980) examined optical properties of polar bear fur –hair is like a drawn quartz tube with irregular inner surface –reflects incident UV towards the skin –light scattering and reflection make the fur look white")
6
Testing hypotheses for adaptations giraffes have long necks to reach leaves on tall trees, right? Wrong.

7
An alternative: neck length evolved through sexual selection Males have necks that are 30-40 cm longer and 1.7 times heavier than females

8
An alternative: neck length evolved through sexual selection Longer-necked males win contests with shorter-necked males and are preferred by females (Pratt and Andersen 1985)
")
9
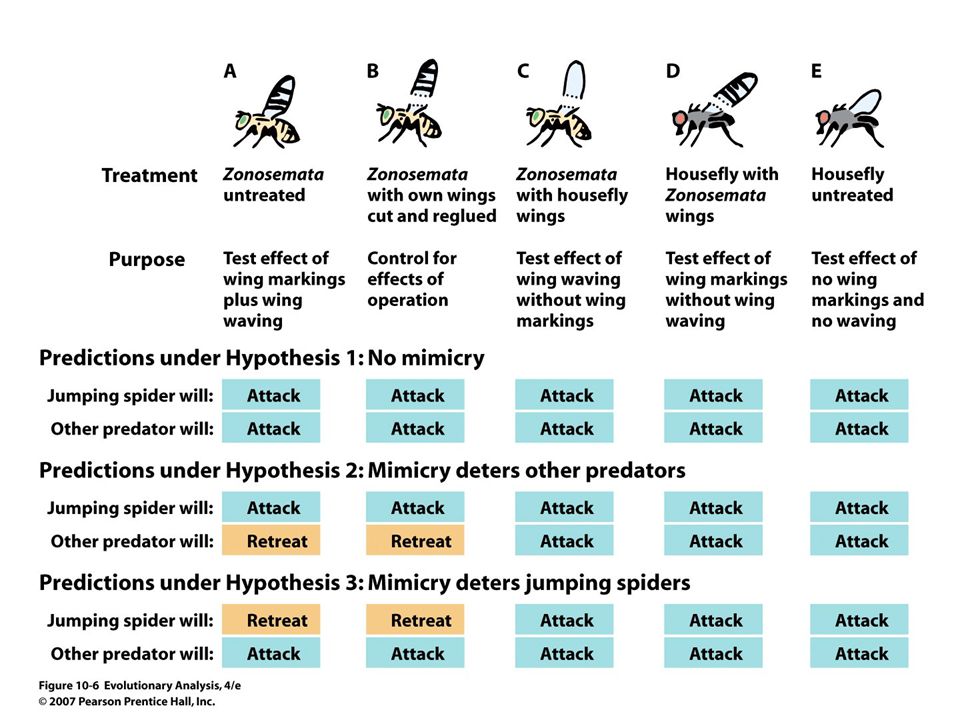
Zonosemata, the prey species Studying adaptation: experimental approaches Are wing markings and waving of Zonosemata adaptations that mimic jumping spider threat display? Phidippus, the predator

10
The hypotheses (1) Zonosemata is not a mimic. Wing markings and display used for courtship? (2) The flies mimic jumping spiders to deter other predators. (3) The flies mimic jumping spider threat displays to deter jumping spiders themselves.
 The flies mimic jumping spiders to deter other predators. (3) The flies mimic jumping spider threat displays to deter jumping spiders themselves..")
12
Results show markings and wing-waving together deter jumping spider predation

13
Mimicry does not deter other predators Tests A, C and E on other spiders, mantises, assassin bugs and whiptail lizards showed no difference in attack times among treatments

14
A sheep in wolf’s clothing!

15
Observational studies: behavioral thermoregulation in reptiles Like all ectotherms, desert iguana internal temperature conforms to the environment They function optimally in a much narrower range of temperatures (35 - 43 C) than found in their environment (15 - 47 C)
 than found in their environment ( C)")
16
Behavioral thermoregulation appears to be adaptive. body temperature “chosen” by iguanas in lab distribution of body temperature in field-caught iguanas from Huey and Kingsolver (1989)
.")
17
Adaptive thermoregulatory behavior in garter snakes In the lab, snakes stay at 28 - 32 C Can they do this in the field, and if so, how? Huey et al. (1989) implanted transmitters to locate snakes and remotely monitor body temperature Western terrestrial form of Thamnophis elegans
 implanted transmitters to locate snakes and remotely monitor body temperature Western terrestrial form of Thamnophis elegans.")
18
Snakes can thermoregulate within a narrow temperature range in the field. critical thermal tolerance limits preferred temperature range

19
Exposed snakes could move from shade to sun to thermoregulate, but are nighttime popsicles! Moving in a burrow is OK but night temperatures are cold

20
Snakes have options during daytime, but overnight the optimal place is under a medium- sized rock

21
Active choice of rocks for nightime cover appears to be adaptive P < 0.05 by chi-square

22
Australian grey-headed flying fox Pteropus poliocephalus The comparative method for studying adaptation In fruit bats and flying foxes (Megachiroptera), is testis size an adaptation to sperm competition?
, is testis size an adaptation to sperm competition")
23
Testis size as an adaptation In bats that roost in larger groups, females mate with more males than in bats with smaller roost group size Sperm competition should increase with group size This may select for larger testes in bats with larger group size

24
Testis size increases with roost group size—appears to support the sperm competition hypothesis. data from Hosken (1998)
.")
25
However, a problem with making comparisons across species is phylogenetic non-independence. Species D,E,F may have larger testes and larger groups because their common ancestor did

26
Then, we want to know: when species diverge from a common ancestor, does the species that evolves a larger group size also evolve larger testes? To proceed, first we need a phylogeny

27
The method of phylogenetically independent contrasts (Felsenstein 1985)
")
28
Independent contrasts applied to the bat data

29
Uncorrected correlation Independent contrasts show that testes mass and group size evolve together from Hosken (1998)
")
30
Phenotypic plasticity Adaptations are nearly always a product of genotype + environment Often, individuals with identical genotypes are phenotypically plastic—they show different phenotypes in different environments Plasticity itself may be adaptive

31
Phenotypic plasticity: phototactic behavior in Daphnia Daphnia is an excellent candidate because females are parthenogenic Identical clones can be tested in different environments De Meester (1996) placed 10 clones in a cylinder and tested phototactic behavior
 placed 10 clones in a cylinder and tested phototactic behavior")
32
Phenotypic plasticity: phototactic behavior in Daphnia A value of +1 means all ten swam towards light, -1 if all swam away De Meester tested clones from each of 3 lakes He also tested whether they changed their behavior in response to fish “scent”

33
(1) Phototactic behavior varies genetically— clones vary in their response (2) Behavior is plastic, particularly in Lake Blankaart
 Phototactic behavior varies genetically— clones vary in their response (2) Behavior is plastic, particularly in Lake Blankaart")
34
(3) Plasticity itself can evolve: clones differ in the extent to which fish induction modifies their behavior
 Plasticity itself can evolve: clones differ in the extent to which fish induction modifies their behavior")
35
(4) Plasticity is adaptive: pronounced in Lake Blankaart (many planktivorous fishes) and absent in Citadelpark (fishless)
 Plasticity is adaptive: pronounced in Lake Blankaart (many planktivorous fishes) and absent in Citadelpark (fishless)")
36
A straightforward hypothesis: fish predation selects for plastic, phototactic behavior To test: De Meester et al. (2001) took sediment cores from 3 depths: representing before, during and after years of intense fish stocking in a manmade pond They hatched resting eggs and scored phototactic behavior of adults
 took sediment cores from 3 depths: representing before, during and after years of intense fish stocking in a manmade pond They hatched resting eggs and scored phototactic behavior of adults.")
37
Daphnia life history from the Daphnia Genomics Consortium

38
Clones hatching from eggs produced during the years of heaviest fish stocking are the most plastic!!

39
An evolutionary trade-off: flower size in Begonia Begonia is monoecious: hermaphrodites with male and female flowers Inflorescence of many flowers of both genders Male flowers (L):pollen reward; Female flowers (R): no nectar
:pollen reward; Female flowers (R): no nectar")
40
Female flowers “mimic” male flowers in color, shape and size: how and why?

41
Schemske and Ågren (1995) H1: selection favors intermediate-sized female flowers resembling the average male flower H2: selection favors larger female flowers resembling the largest (most attractive) male flowers
 H1: selection favors intermediate-sized female flowers resembling the average male flower H2: selection favors larger female flowers resembling the largest (most attractive) male flowers")
42
Their experiment... Equal numbers of small, medium and large artificial flowers arrayed in a natural forest population Approaches Visits (pollinator lands) H2 is correct: there is directional selection for larger male flowers And results...
 H2 is correct: there is directional selection for larger male flowers And results....")
43
So why are female flowers intermediate in size? An evolutionary trade- off: the larger the flower, the smaller the number of flowers that can be maintained in a cluster (inflorescence) A trade-off hypothesis: opposing forces of selection for flower size and inflorescence size
 A trade-off hypothesis: opposing forces of selection for flower size and inflorescence size.")
44
Evolutionary constraints on adaptation: flower color change in Fuchsia F. excorticata: a flowering bird- pollinated tree, endemic to New Zealand Has showy flowers that are green when exporting and receiving pollen, then turn and stay red for ~ 5 days

45
from Delph and Lively (1989) pollination occurs before day 8, but red flowers remain for ~5 days more exporting pollen receiving pollen
 pollination occurs before day 8, but red flowers remain for ~5 days more exporting pollen receiving pollen")
46
Why do flowers turn red? Color change is a cue to pollinators to avoid flowers with no nectar reward Pollinator “efficiency” benefits flowers –inviable pollen is not transferred –nonreceptive stigmas do not receive viable pollen

47
But why does Fuchsia retain red flowers so long? Dropping flowers would be a better signal to pollinators It would be energetically cheaper

48
An adaptive hypothesis H1: red flowers attract birds and receptive green flowers on the same plant receive more pollen than without color change –Delph and Lively removed red flowers from some trees and not from others –Trees with and without red flowers: no difference in pollen deposited on green flowers

49
A “constraint” hypothesis H2: Fuchsia is physiologically constrained to retain red flowers long after pollination is complete –Delph and Lively removed hand-pollinated flowers –They dissected 10 flowers at a time to see whether pollen tubes had reached the ovary

50
Their results Fuschia is constrained to retain its flowers. It was selected to change flower color to provide a reliable pollinator cue.

51
fertilization complete abcission layer complete exporting pollen receiving pollen

Similar presentations