Download presentation
Presentation is loading. Please wait.

3
Eduard Buchner (1860-1917) 1897 found fermentation in broken yeast cells 1907 Nobel Prize in Chemistry
 1897 found fermentation in broken yeast cells 1907 Nobel Prize in Chemistry")
4
The whole pathway in yeast and muscle cell were elucidated by Arthur Harden 1865-1940

8
Glycolysis Glycolysis is an almost universal central pathway of glucose catabolism, the pathway with the largest flux of carbon in most cells. In some mammalian tissues (erythrocytes, renal medulla, brain, sperm), the glycolytic breakdown of glucose is the sole source of metabolic energy.
, the glycolytic breakdown of glucose is the sole source of metabolic energy..")
9
Glycolysis Some of the starch-storing tissues, like potato tubers, and some aquatic plants derive most of their energy from glycolysis. Many anaerobic microorganisms are entirely dependent on glycolysis.

10
1. phosphorylation of glucose

11
2. Isomerization of glucose 6- phosphate

12
Phosphohexose isomerase reaction by an active-site His residue Glu

13
3. Phosphorylation of fructose 6- phosphate: the first committed step in glycolysis

14
PFK-1 is named so because there is another enzyme catalyzes a similar reaction

15
In some bacteria, protists and (all) plants, a pyrophosphate-dependent phosphofructokinase (PFP) also catalyzes this reaction in a reversible way
 plants, a pyrophosphate-dependent phosphofructokinase (PFP) also catalyzes this reaction in a reversible way")
16
4. Cleavage of fructose 1,6- bisphosphate

17
Class I aldolases form Schiff base intermediate during sugar cleavage reaction Class I aldolases were found in animals and plants. Class II aldolases (fungi and bacteria) do not form the Schiff base and require a zinc ion to catalyze reaction.
 do not form the Schiff base and require a zinc ion to catalyze reaction..")
18
5. Interconversion of the triose phosphate

19
Dihydroxyacetone phosphate and glyceraldehyde 3-phosphate become indistinguishable after triose phosphate isomerase reaction

20
6. Oxidation of glyceraldehyde 3- phosphate to 1,3-bisphosphoglycerate

21
The glyceraldehyde 3-phosphate dehydrogenase reaction hemiacetal Heavy metal ion such as Hg 2+ will react with Cys residue, hence irreversibly inhibits the enzyme.

22
7. Phosphoryl transfer from 1,3- bisphosphoglycerate to ADP

23
Glyceraldehyde 3-phosphate dehydrogenase and Phosphoglycerate kinase are coupled in vivo Glyceraldehyde 3-phosphate dehydrogenase catalyzes an endergonic reaction while phosphoglycerate kinase catalyzes an exergonic reaction. When these two reactions are coupled (which happens in vivo), the overall reaction is exergonic.
, the overall reaction is exergonic..")
24
Substrate-level phosphorylation soluble enzymes chemical intermediates Respiration-linked phosphorylation Photophosphorylation membrane-bound enzymes transmembrane gradients of protons The formation of ATP by phosphoryl group transfer from a substrate is referred to as a substrate-level phosphorylation

25
8. Conversion of 3- phosphoglycerate to 2- phosphoglycerate

26
The phosphoglycerate mutase reaction

27
2,3-Bisphosphoglycerate (BPG) The concentration of BPG is usually low in most of the tissues except erythrocytes (up to 5 mM). Function of BPG in erythrocytes is to regulate the affinity of hemoglobulin to O 2.

28
9. Dehydration of 2- phosphoglycerate to phosphoenolpyruvate

29
10. Transfer of the phosphoryl group from phosphoenolpyruvate to ADP

31
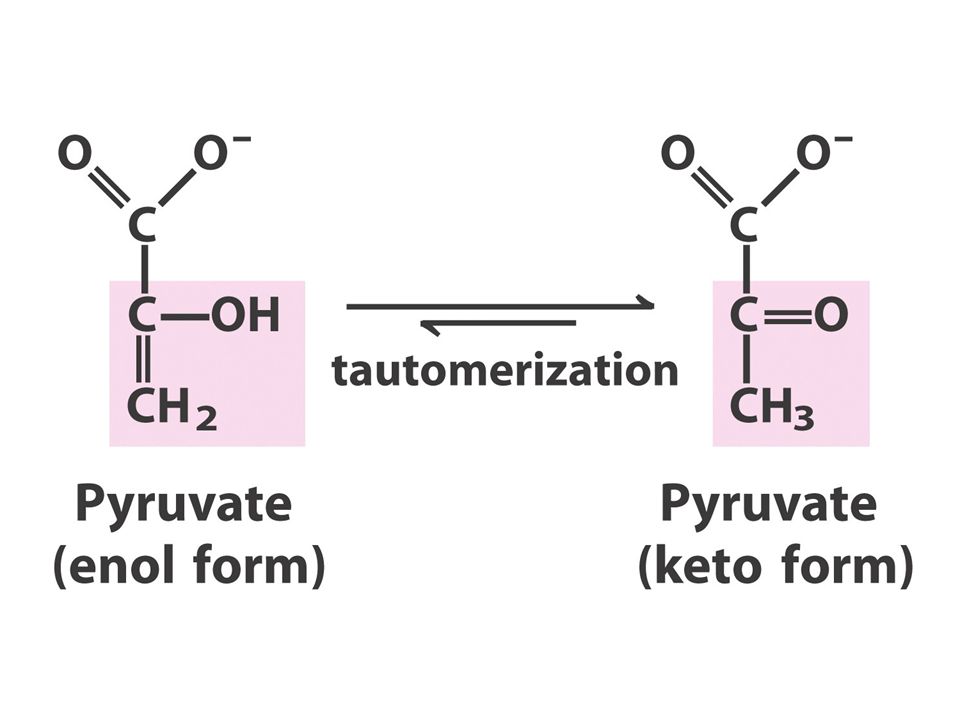
Glucose + 2ATP + 2NAD + + 4ADP + 2Pi 2 pyruvate + 2ADP + 2NADH + 2H + + 4ATP + 2H 2 O Glucose + 2ADP + 2NAD + + 2Pi 2 pyruvate + 2ATP + 2NADH + 2H + 在有氧狀況下,產生的 NADH 很快就被送 到 mitochondria 中用來合成 ATP

32
NAD + (nicotinamide adenine dinucleotide) is the active form of niacin
 is the active form of niacin")
34
Niacin is the common name for nicotinamide and nicotinic acid. Nicotinic acid is the common precursor for NAD + and NADP + biosynthesis in cytosol. Niacin

35
Functions of NAD + and NADP + Both NAD + and NADP + are coenzymes for many dehydrogenases in cytosol and mitochondria NAD + is involved in oxidoreduction reactions in oxidative pathways. NADP + is involved mostly in reductive biosynthesis.

37
Weight loss, digestive disorders, dermatitis, dementia Niacin deficiency: pellagra

38
Niacin deficiency Because niacin is present in most of the food and NAD + can also be produced from tryptophan (60 grams of trptophan 1 gram of NAD + ), so it is not often to observe niacin deficiency. However, niacin deficiency can still be observed in areas where maize is the main carbohydrate source because maize only contain niacytin, a bound unavailable form of niacin. Pre-treated maize with base will release the niacin from niacytin.

39
Niacin deficiency Areas where sorghum is the main carbohydrate source will also observe niacin deficiency if niacin uptake is not being watched carefully. Sorghum contains large amount of leucine, which will inhibit quinolinate phosphoribosyl transferase (QPRT), an enzyme involved in NAD + biosynthesis from tryptophan. Vitamin B6 deficiency can also lead to niacin deficiency because pyridoxal phosphate is a coenzyme in NAD + biosynthesis from tryptophan.
, an enzyme involved in NAD + biosynthesis from tryptophan. Vitamin B6 deficiency can also lead to niacin deficiency because pyridoxal phosphate is a coenzyme in NAD + biosynthesis from tryptophan..")
40
ISONIAZID A Commonly Used Medication for HIV & AIDS Patients Drug: ISONIAZID Classification: Antimycobacterial Indication: Infection with, or disease from, mycobacterium tuberculosis

41
Feeder pathways for glycolysis

42
Glycogen and starch are degraded by phosphorolysis Glycogen and starch can be mobilized for use by a phosphorolytic reaction catalyzed by glycogen/starch phosphorylase. This enzyme catalyze an attack by Pi on the ( 1 4) glycosidic linkage from the nonreducing end, generating glucose 1- phosphate and a polymer one glucose unit shorter.
 glycosidic linkage from the nonreducing end, generating glucose 1- phosphate and a polymer one glucose unit shorter..")
44
Branch point ( 1 6) is removed by debranching enzyme
 is removed by debranching enzyme")
45
Glucose 1-phosphate is converted to G-6-P by phosphoglucomutase by the same mechanism observed in phosphoglycerate mutase reaction

46
Digestion of dietary polysaccharides Digestion begins in the mouth with salivary - amylase hydrolyze (attacking by water) the internal glycosidic linkages. Salivary -amylase is then inactivated by gastric juice; however pancreatic -amylase will take its place at small intestine. The products are maltose, maltotriose, and limit dextrins (fragments of amylopectin containing 1 6 branch points.

47
Digestion of dietary disaccharides Disaccharides must be hydrolyzed to monosaccharides before entering cells. Dextrin + nH 2 O n D-glucose Maltose + H 2 O 2 D-glucose Lactose + H 2 O D-galactose + D-glucose Sucrose + H 2 O D-fructose + D-glucose Trehalose + H 2 O 2 D-glucose dextrinase maltase lactase sucrase trehalase

48
Lactose intolerance Lactose intolerance is due to the disappearance after childhood of most or all of the lactase activity of the intestinal cells.

49
Lactose intolerance Undigested lactose will be converted to toxic products by bacteria in large intestine, causing abdominal cramps and diarrhea.

50
Fructose metabolism in muscle and kidney

51
Fructose metabolism in liver In liver, the enzyme fructokinase catalyze the phosphorylation of fructose to form fructose 1-phosphate. Triose phosphate isomerase

52
Galactose metabolism Galactose is phosphorylated by galactokinase in the liver. Then galactose 1-phosphate is converted to glucose 1-phosphate by a series of reactions.

53
Galactose metabolism The conversion of galactose 1-P to glucose 1-P (epimerization) requires uridine diphosphate (UDP) as a coenzyme-like carrier of hexose groups.
 requires uridine diphosphate (UDP) as a coenzyme-like carrier of hexose groups.")
54
Galactosemia inability to metabolize galactose due to lack of 1. UDP-glucose galactose 1-phosphate uridylyltransferase (classical galactosemia) 2. UDP-glucose 4-epimerase 3. Galactokinase Among these, deficiency of either 1 or 2 is more severe (1 is the most severe).
 2. UDP-glucose 4-epimerase 3. Galactokinase Among these, deficiency of either 1 or 2 is more severe (1 is the most severe)..")
55
Galactosemia Deficiency of transferase (or epimerase) will result in poor growth, speech abnormality, mental deficiency, and (fatal) liver damage even when galactose is withheld from the diet.
 will result in poor growth, speech abnormality, mental deficiency, and (fatal) liver damage even when galactose is withheld from the diet.")
56
Galactosemia patients develop cataracts by deposition of galactitol in the lens

57
Mannose + ATP mannose 6-phosphate hexokinase +ADP mannose 6-phosphate fructose 6-phosphate phosphomannose isomerase Mannose metabolism

58
Fermentation Fermentation is referring to the process when no oxygen is consumed or no change in the concentration of NAD + or NADH during energy extraction.

59
Fermentation Under hypoxic conditions, oxidative phosphorylation will be the first to stop. Then citric acid cycle will come to a halt due to inhibition effect from NADH. As a result, glycolysis will be the only metabolic pathway that is available to energy production during hypoxia.

60
Fermenation However, the oxidation of glyceraldehyde 3- phosphate consumes NAD + that will not be regenerated under hypoxic condition because oxidative phosphorylation is not available.

61
The purpose of fermentation is to regenerate NAD + In order to continue regenerating NAD +, cells come up a strategy. During fermentation, NAD + is regenerated during the reduction of pyruvate, the product of glycolysis.

62
Lactate fermentation glycolysis

63
Lactate is recycled in the liver (Cori cycle)
")
64
Carl and Gerty Cori, 1947 Nobel Prize in Physiology and Medicine

65
Lactate fermentation only happened in larger animals Most small vertebrates and moderate size running animals have circulatory systems that can carry oxygen to their muscles fast enough to avoid having to use muscle glycogen anaerobically.

66
http://www.mountain-research.org/CV/coelacanth.jpg http://www.anac.8m.net/Images/coelacanth.jpg Deep sea fish (below 4,000 m or more) coelacanth uses anaerobic metabolism exclusively. The lactate produced is excreted directly. Some marine vertebrates can do ethanol fermentation.

67
Ethanol fermentation Yeast and other microorganisms ferment glucose to ethanol and CO 2. Pyruvate is first decarboxylated by pyruvate decarboxylase, which is absent in vertebrate tissues and in other organisms that carry out lactic acid fermentation. Acetaldehyde is the product of this reaction.

68
Pyruvate decarboxylase The decarboxylation of pyruvate by pyruvate decarboxylase produces CO 2, which is the reason why champagne is bubbling.

69
Thiamine pyrophosphate (TPP) is the coenzyme of pyruvate decarboxylase Thiamine pyrophosphate is derived from vitamin B1 (thiamine). Lack of vitamine B1 will lead to beriberi (edema, pain, paralysis, death).
..")
70
TPP plays an important role in the cleavage of bonds adjacent to a carbonyl group. The thiazolium ring of TPP acts as an “electron sink” to facilitates decarboxylation reaction.

71
Alcohol dehydrogenase catalyze the second step of ethanol fermentation Alcohol dehydrogeanse reduces acetaldehyde, producing NAD + and ethanol. This enzyme is present in many organisms that metabolize ethanol, including human.

72
Fermentation has commercial values Bacteria like Lactobacillus bulgaricus (yogurt) and Propionibacterium freudenreichii (swiss cheese) ferments milk to produce lactic acid or propionic acid and CO 2.
 and Propionibacterium freudenreichii (swiss cheese) ferments milk to produce lactic acid or propionic acid and CO 2.")
73
Dr. Chaim Weizmann 1874-1952 First President of Israel Found butanol and acetone fermentation in Clostridium acetobutyricum

74
Industrial fermentation is done in huge close vats Fermentors are huge closed vats in which temperature and access to air are adjusted to favor the multiplication of the desired microorganism. Some even immobilize the cells in an inert support so no effort is required to separate microorganisms from products after fermentation is completed.

Similar presentations












 1. Why gluconeogenesis?>")


 Lactate or ethanol (anaerobic) (2) Acetyl CoA (aerobic) Acetyl CoA.>")





 polyhydroxy aldehydes. b) polyhydroxy ketones. c) polyhydroxy acids. d) polyhydroxy alcohols.>")
 to 2 pyruvate (3C) – produces: 4 ATP & 2 NADH – consumes: 2 ATP – net: 2 ATP & 2 NADH Overview DHAP = dihydroxyacetone.>")



 October 17, 2003 Haining Zhu Dept. of Molecular.>")
