Download presentation
Presentation is loading. Please wait.

1
the phage lambda ( ) A phage (from 'bacteria' and Greek phagein ( , 'to eat') is a virus that infects and sometimes lyses bacteria. Like viruses that infect eukaryotes, phages have several aspects. Typically, they consist of a hull and the enclosed genetic material. The genome is made of ds DNA of 5 to 650 kbp. The dimensions of a phage is on the order of 24 to 200 nm. Several phages have a tail, which is used to inject the genetic material into the host. Phages can be found in many reservoirs populated by bacteria such as soil, the intestine of animals etc. One of the densest natural sources for phages is seawater, where up to 2.5x108 virions per milliliter have been found. -------------------------- hull: el casco.

2
Lysis and Lysogeny The lambda phage is adorbed on the lamb receptor. This receptor is only expressed when the bacteria are grown in the presence of maltose. Such a bacteriuml is known as “competent”. Lysis: multiplication. Most phages take over the machinery of the host cell to produce a large number of viral particles. This result in the lysis of the host cell, which releases the newborn phages. 2. Lysogeny: rest. In some infections, a phage stays silent within the cell. This silent phage can go lytic under certain conditions. a lysogenic phage is called a prophage (integrated into the chromosome) Lysogenic bacteria have immunity against further infection The prophage can be induced and is excised from the bacterial genome
 Lysogenic bacteria have immunity against further infection The prophage can be induced and is excised from the bacterial genome.")
3
Lysis and lysogeny are controlled by the proteins encoded by cro and cI genes. The lambda phage will remain in the lysogenic state if cI proteins predominate, but will be transformed into the lytic cycle if Cro proteins predominate cI genes transcription and translation regulate lysis/lysogeny

4
The cI dimer may bind to any of three operators, OR1, OR2, and OR3, in the order OR1 > OR2 > OR3. Binding of a cI dimer to OR1 enhances binding of a second cI dimer to OR2: thus, OR1 and OR2 are almost always simultaneously occupied by cI dimer. This does not increase the affinity between cI dimer and OR3, which will be occupied only when the cI dimer concentration is high. (a) In the absence of cI proteins, the cro gene may be transcribed ---> lysis (b) In the presence of cI proteins, only the cI gene may be transcribed --->lysogeny (c) At high concentration of cI, transcriptions of both genes are repressed ---> lysogeny When the host DNA is damaged (e.g., under UV irradiation), the cI protein may be cleaved by certain protease promoted by the RecA protein. Cleaved cI proteins cannot bind to the operators. Thus, the Cro proteins can be produced to transform the lambda phage into the lytic cycle.
 In the absence of cI proteins, the cro gene may be transcribed ---> lysis (b) In the presence of cI proteins, only the cI gene may be transcribed --->lysogeny (c) At high concentration of cI, transcriptions of both genes are repressed ---> lysogeny When the host DNA is damaged (e.g., under UV irradiation), the cI protein may be cleaved by certain protease promoted by the RecA protein. Cleaved cI proteins cannot bind to the operators. Thus, the Cro proteins can be produced to transform the lambda phage into the lytic cycle..")
5
Lytic or Virulent Phages

9
the phage genome

10
modes of replication: Cairn and rolling circle

11
1. Definition A lysogenic phage is a phage that enters a quiescent state in the cell. In this quiescent only the cI gene is transcribed: the phage genome exists in a repressed state. In this repressed state, the phage DNA is called a prophage A prophage don’t have the potential to produce other phages. In most cases the phage DNA actually integrates into the host chromosome and is replicated along with the host chromosome and passed on to the daughter cells. The cell harboring a prophage is not adversely affected by the presence of the prophage and the lysogenic state may persist indefinitely. The cell harboring a prophage is termed a lysogen. Lysogenic and integration into the bacterial genome.

12
2. Events Leading to Lysogeny - The Prototype Phage: Lambda a. Circularization of the phage chromosome - Lambda DNA is a double stranded linear molecule with small single stranded regions at the 5' ends. These single stranded ends are complementary (cohesive ends) so that they can base pair and produce a circular molecule. In the cell the free ends of the circle can be ligated to form a covalently closed circle as illustrated below.
 so that they can base pair and produce a circular molecule. In the cell the free ends of the circle can be ligated to form a covalently closed circle as illustrated below..")
13
b. Site-specific recombination In phage lambda, the integration site is known as att P, in E. coli the site is att B. The att sites contain the binding sites for the proteins that mediate lambda recombination. The integration reaction (att B x att P) is mediated by the proteins integrase (Int) and host integration factor (IHF).
 is mediated by the proteins integrase (Int) and host integration factor (IHF)..")
14
integration: When integration occurs, two new sites are created, att L and att R, flanking the integrated prophage, with no loss of DNA sequence. All the att sites are alike in that they contain a 15-bp recognition sequence for the recombinase (integrase). excision: When att L x att R recombine (mediated by the proteins integrase and host integration factor and excisionase [Xis]), the lambda -DNA is excised from the E. coli genome, recreating the att B site in E. coli and the att P site in lambda.
. excision: When att L x att R recombine (mediated by the proteins integrase and host integration factor and excisionase [Xis]), the lambda -DNA is excised from the E. coli genome, recreating the att B site in E. coli and the att P site in lambda..")
15
c. Repression of the phage genome - A phage coded protein, a repressor, is made which binds to a particular site on the phage DNA, called the operator, and shuts off transcription of most phage genes EXCEPT the repressor gene. The result is a stable repressed phage genome which is integrated into the host chromosome. Each temperate phage will only repress its own DNA and not that from other phage, so that repression is very specific (immunity to superinfection with the same phage). http://pathmicro.med.sc.edu/mayer/phage.htm http://pathmicro.med.sc.edu/mayer/phage.htm d. Lysogenic conversion - When a cell becomes lysogenized, occasionally extra genes carried by the phage get expressed in the cell. These genes can change the properties of the bacterial cell. This process is called lysogenic or phage conversion. This can be of significance clinically. e.g. Lysogenic phages have been shown to carry genes that can modify the Salmonella O antigen, which is one of the major antigens to which the immune response is directed. Toxin production by Corynebacterium diphtheriae is mediated by a gene carried by a phage. Only those strain that have been converted by lysogeny are pathogenic
. d. Lysogenic conversion - When a cell becomes lysogenized, occasionally extra genes carried by the phage get expressed in the cell. These genes can change the properties of the bacterial cell. This process is called lysogenic or phage conversion. This can be of significance clinically. e.g. Lysogenic phages have been shown to carry genes that can modify the Salmonella O antigen, which is one of the major antigens to which the immune response is directed. Toxin production by Corynebacterium diphtheriae is mediated by a gene carried by a phage. Only those strain that have been converted by lysogeny are pathogenic.")
16
Gatway system (Invitrogen): This reaction involves an entry clone, a destination vector, and a mix which contains the recombination proteins necessary for excision and incision (incubation time: one hour).
: This reaction involves an entry clone, a destination vector, and a mix which contains the recombination proteins necessary for excision and incision (incubation time: one hour).")
19
The Lambda EMBL3 vector (Stratagen, Promega) is a genomic replacement lambda phage vector capable of accepting BamH I-compatible fragments (Sau3A I, Mbo I, Bgl II,or BamH I), ranging in size from 9 to 23 kb (Figure above). The arms are prepared by double digestion with Bam H I and EcoR I followed by a selective precipitation which removes the small BamH I/EcoR I linker thatseparate the arms from the stuffer fragment. Because this treatment leavesthe arms with BamH I ends and the stuffer fragment with EcoR I ends, there is no need to physically separate them. Target DNA cloned into theBamH I sites of EMBL3 may be removed by digestion with Sal I. Thered and gam genes are located on the stuffer fragment; therefore, WT phagecannot grow on the P2 lysogenic strain. When the stuffer fragment is replaced by an insert, the recombinant phage becomes red–/gam–, and the phage is ableto grow on the P2 lysogenic strain. Therefore, by plating the library on P2 lysogenic strain, only recombinant phages are allowed to grow. The Lambda EMBL3 vector

20
packaging the phage recombinant DNA Here are the genes coding for the head and the tail of the phage Protein coded by the gene A: The protein A is necessary to fill the head with “one” genome and to cut the concatemer when “one genome” is packaged into the head. Without A, the heads stay empty. Protein coded by the gene E: Without E there is no heads. Protein E is one of the main constituent of the phage head.

21
(the method described below is only an example, there other methods) The packaging uses two kinds of bacteria, each infected by a lysogenic phages: the first phage has a mutation in protein A: in its lytic form, it is unable to package DNA into a head and to cut the concatemer at the cos sites the second phage has a mutation in protein E: in its lytic form,it doesn’t buit a head. the head parts are disseminated through the cytoplasm. Both phage are maintained in the lysogenic form by a thermosensible cI (the protéin acts at 32°. Above, it is destroyed and the phage becomes lytic. The two kind of bacteria show another peculiarity. Their DNA have not attB sequences. This mutation doesn’t allow this DNA to be packaged. An extract of both phages is mix with the recombinant ADN at 45° C. The recovery is low, between 0.05 and 0.5 %. The method can be used with cosmid DNA.

35
Adeno-associated-viruses (AAVs) The 5 serotypes of the human AAVs belong to the genus Dependovirus. AAVs are similar to parvoviruses. They are not replicated by any cells. To replicate them, the host cell has also to be infected by helpers viruses. The sytem described above is sold by Stratagen. It is not dangerous for the people in the lab. It contains no viruses but: 3 plasmids (presented above) an eukaryotic cell able to produce them (AVV-293 cell), a modified HEK-293 cell.
 an eukaryotic cell able to produce them (AVV-293 cell), a modified HEK-293 cell..")
36
A no helper virus system (eliminating the requirement for live helpervirus ): Stratagene’s AAV Helper-Free System allows the production of infectious recombinant hAAV-2 virions without the use of any helper virus. This system takes advantage of the identification of the 4 specific adenovirus genes (e2A, e4, and va RNA) that mediate AAV replication and the demonstration that some of these gene can be introduced into a modified host cell (AAV-293) by transfection through the helper plasmid: 3 out of 4 adenovirus gene whose products are required for the production of infective AAV particles are supplied on the plasmid pHelper (i.e. e2A, e4, and va RNA genes). this plasmid is co-transfected into cells. The remaining adenoviral gene (e1) is supplied by the AAV-293 host cells. A plasmid to get more room for the insert this plasmid bears the genes re and cap. AAV-293cells are HEK293-derived cells with improved adeno-associated virus production capabilities.
 that mediate AAV replication and the demonstration that some of these gene can be introduced into a modified host cell (AAV-293) by transfection through the helper plasmid: 3 out of 4 adenovirus gene whose products are required for the production of infective AAV particles are supplied on the plasmid pHelper (i.e. e2A, e4, and va RNA genes). this plasmid is co-transfected into cells. The remaining adenoviral gene (e1) is supplied by the AAV-293 host cells. A plasmid to get more room for the insert this plasmid bears the genes re and cap. AAV-293cells are HEK293-derived cells with improved adeno-associated virus production capabilities..")
37
ITR: inverted repeat sequences (secuencias repetitivas inversadas) Not I: site for the restriction enzyme Not 1. pAAV-MCS (Multiple Cloning Site Region, nt 1319-1394) Cla I EcoR I BamH I Xba IHinc II Hind III Xho I Bgl II atcgattgaattccccggggatcctctagagtcgacctgcagaagcttgcctcgagcagcgctgctcgagagatct
 Cla I EcoR I BamH I Xba IHinc II Hind III Xho I Bgl II atcgattgaattccccggggatcctctagagtcgacctgcagaagcttgcctcgagcagcgctgctcgagagatct.")
39
pAAV-MCS (Multiple Cloning Site Region, nt 1319-1394) Cla I EcoR I BamH I Xba IHinc II Hind III Xho I Bgl II atcgattgaattccccggggatcctctagagtcgacctgcagaagcttgcctcgagcagcgctgctcgagagatct
 Cla I EcoR I BamH I Xba IHinc II Hind III Xho I Bgl II atcgattgaattccccggggatcctctagagtcgacctgcagaagcttgcctcgagcagcgctgctcgagagatct")
40
rep and cap genes are on another plasmid creating a 3 kb extra space. Rep and Cap proteins are implicated in replication and capside formation respectively. El genóma salvaje AAV-2 contiene los genes rep y cap que codifican Rep y CAP, proteînas implicadas respectivamente en la replicaciôn y en la formaciôn de la câpside. Esos genes contienen dos secuencias terminales repetitivas (ITRs) que contienen los elementos cis necesarios para la replicaciôn y el empaquetamiento. En el sistema sin helper (helper free), se han metido rep y cap en un plâsmido. Al sacar êsos genes, se crea un espacio de 3kb que son los utilizados para ahî meter el inserto.
 que contienen los elementos cis necesarios para la replicaciôn y el empaquetamiento. En el sistema sin helper (helper free), se han metido rep y cap en un plâsmido. Al sacar êsos genes, se crea un espacio de 3kb que son los utilizados para ahî meter el inserto..")
41
The “helper” plamid contains 3 genes that force the host cell to replicate the virus producing particles able to infect the target cell. El plásmido "Helper” que contiene tres de los genes (E2A, E4 y VA RNA) que obligan a la célula huésped a replicar el virus y a producir partículas capáces de infectar la célula target.
 que obligan a la célula huésped a replicar el virus y a producir partículas capáces de infectar la célula target..")
42
4. Las cêlulas huêsped, AVV-293 Esas cêlulas pueden proveer a otros genes virales, notablemente el gen E1 de los adenovirus.

43
examples Human immunoglobulin inhibits liver transduction by AAV vectors at low AAV neutralizing titers in SCID mice Ciaran D. Scallan, Haiyan Jiang, Tongyao Liu, Susannah Patarroyo-White, Jurg M. Sommer, Shangzhen Zhou, Linda B. Couto,and Glenn F. Pierce Blood, 1 March 2006 Volume 107, Number 5

44
Adeno-associated virus (AAV) vectors have reproducibly delivered transgenes in animal models, resulting in successful treatment and cure of numerous clinically relevant diseases. These studies have demonstrated sustained transgene expression without toxicity in mice, rats, dogs, and nonhuman primates. In addition, severa lclinical trials using AAV2 vectors have shown excellent safety profiles; however, clear evidence of therapeutic efficacy has not yet been Achieved. A major challenge for AAV-mediated genetherapy is pre-existing AAV-specific antibodies prevalent in the human population resulting from prior natural infections. Theeffect of these pre-existing antibodies on AAV gene transfer variesamong individuals, depending on actual neutralizing titers, thedelivery route, and target organ. Compartments that have limited access to systemically circulating antibodies, such as brain and skeletal muscle, have been transduced successfully in the presenceof high circulating Neutralizing titers. Intravascular delivery to target organs such as the liver will likely be more Problematic because the circulating antibodies can rapidly bind to antigens at an average Kd of 10 7 to 10 11 M.

45
Adenoviruses Human serotype 5 adenoviruses are also used to introduce in mammalian cells, genes we want to express. Adenoviruses don’t need cell in a reproductive phase. Serotype 5,which is rendered replication defective by deletion of the genes E1 and E3. This deletion gives an insertion space of 7.5 kb E1 is essential for the assembly of infectious virus particles and is complemented E3 encodes proteins involved in evading host immunity. As E1 is necessary, we complement it by modified HEK 293 cell that express E1 (AD-293). As the virus posses a very small number of restriction sites, the gene of interest is first inserted in a shuttle vector. Once the gene is inserted in the shuttle, it is inserted in pEasy by recombibation
. As the virus posses a very small number of restriction sites, the gene of interest is first inserted in a shuttle vector. Once the gene is inserted in the shuttle, it is inserted in pEasy by recombibation.")
46
Because the adenovirus genome is large (36 kb) and contains few restriction sites, the gene of interest is first inserted into a shuttle vector and then transferred into the adenovirus genome by means of homologous recombination. Employing the efficient homologous recombination machinery in E. coli, a recombinant adenovirus is producedby a double-recombination event between cotransformed adenoviral backbone plasmid vector, pAdEasy-1, and a shuttle vector carrying the gene of interest. This eliminates the need to manipulate the large adenovirus DNA molecule in vitro (in restriction and ligation reactions).
..")
47
ES = encapsidatoion site

51
practice --------------------------- balance: (aparato) balanza sterile petri dishes: placas Petri estériles 5 tubes with metal caps: 5 tubos de ensayo con tapas de metal. an "Erlenmeyer" flask with a gauze cap: un Erlenmeyer provisto de buchón estéril. Gilson pipettes on a white rack and tips: pipetas Gilson y puntas spatula: paleta, espátula

52
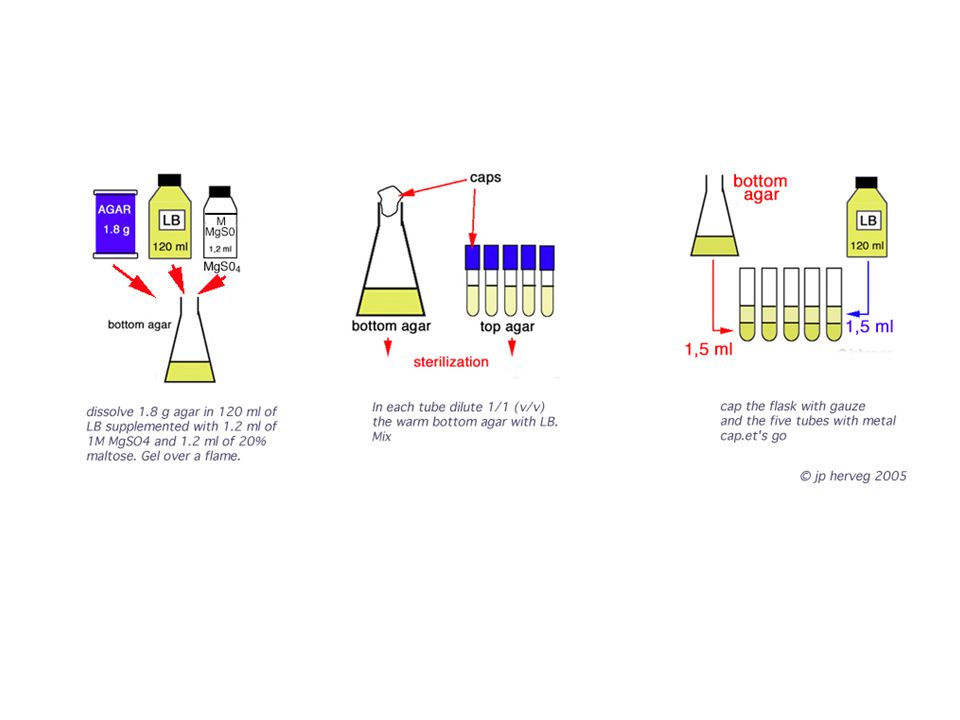
1. Select the material you'll use today 2. Prepare the petri dishes with two agar layers: bottom and top bottom agar 1,5 g % of agar in LB medium (LB = Luria-Bertoni) top agar: a 1/1 dilution of botoom agar --------------------------- 1. Selecciona el material a usar. 2. Prepara placas de agar con dos capas : 5 sterile petri dishes: 5 placas Petri estériles 5 tubes with metal caps: 5 tubos de ensayo con tapas de metal. Bottom and top agar: la capa inferior (agar de base) y la capa superior.
 top agar: a 1/1 dilution of botoom agar Selecciona el material a usar. 2. Prepara placas de agar con dos capas : 5 sterile petri dishes: 5 placas Petri estériles 5 tubes with metal caps: 5 tubos de ensayo con tapas de metal. Bottom and top agar: la capa inferior (agar de base) y la capa superior..")
53
3. Sterilize both agar bottom agar in an erlenmeyer top agar in tubes. 4. Pour Bottom agar in petri dishes (using the laminary flow hood) 5. Place top agar tubes at 47° while you are infecting the competent E. coli. --------------------------------- 3. Estériliza en una olla de presión: el agar de base (en erlenmeyer) y el agar superior (en tubos de ensayo). 4. Vierte el agar de base en las placas (en la cámara de flujo laminar) 5. Manten el agar superior a 47 °C mientras infecta la E. coli. competente
 5. Place top agar tubes at 47° while you are infecting the competent E. coli Estériliza en una olla de presión: el agar de base (en erlenmeyer) y el agar superior (en tubos de ensayo). 4. Vierte el agar de base en las placas (en la cámara de flujo laminar) 5. Manten el agar superior a 47 °C mientras infecta la E. coli. competente.")
54
6. Mix the infected E coli with top agar, mix, and pour on top of bottom agar. 7. Let cool down in the incubator: now you have two layers -------------------------- 6. Siembra la E. coli infectada (mezclada con el agar superior a 47°C) y viertelo sobre el agar de base. 7. Dejalo enfriar antes de meterlo en la estufa.
 y viertelo sobre el agar de base. 7. Dejalo enfriar antes de meterlo en la estufa..")
55
1. Select the material you'll use today 2. Prepare the petri dishes with two agar layers: bottom and top bottom agar 1,5 g % of agar in LB medium top agar: a 1/1 dilution of bottom agar

56
3. Sterilize both agar bottom agar in an erlenmeyer top agar in tubes.

57
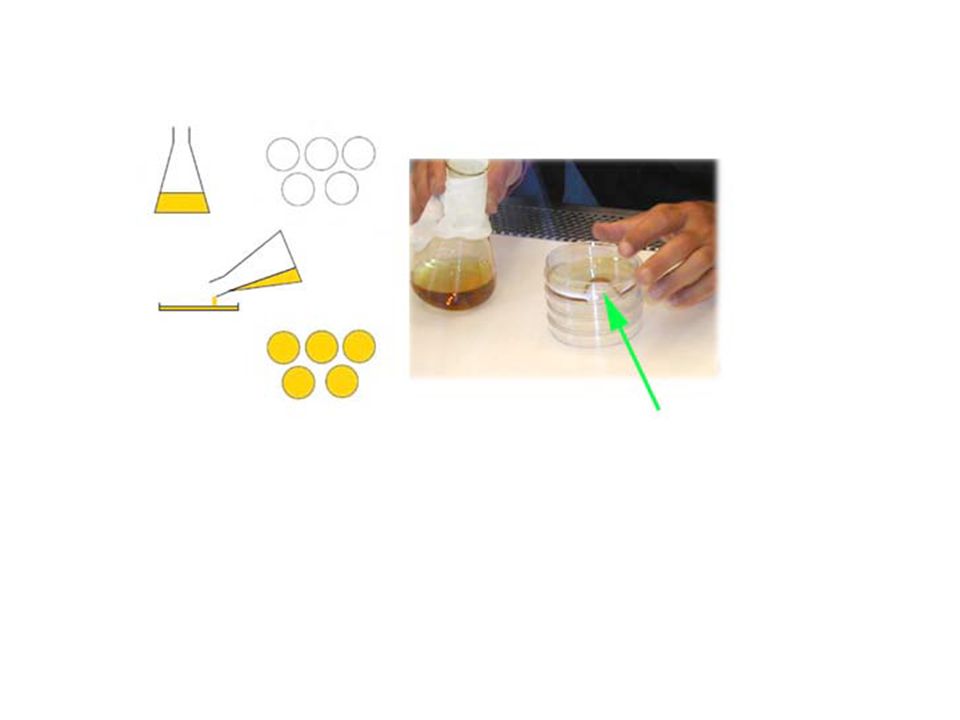
4. Pour Bottom agar in petri dishes (using the laminary flow hood)
")
58
5. Place top agar tubes at 47° while you are infecting the competent E. coli.

59
6. Mix the infected E coli with top agar, mix, and pour on top of bottom agar

Similar presentations





































>")
 Lecture 11 Biotechnology (Text Chapters: 10.15-10.17; 31.1-31.10)>")
 Viruses that target bacteria Virus defining characteristics: parasitic entities Nucleic acid molecules protected by protein.>")





