Download presentation
Presentation is loading. Please wait.

1
Chapter 21. Development of Multicellular Organisms

2
Frog Development

3
Developmental process against the 2nd law of Thermodynamics ?
Developmental processes cause -Increase of orderliness? -Decrease of entropy?

4
The 2nd Law of thermodynamics

5
Information is -ΔG Maxwell’s demon
Information is -ΔG

6
The Genome is The Maxwell’s demon
Life is open system Life has information Life consumes –ΔG, (Erwin Schrodinger, )
")
7
Selective gene expression control four processes by which the embryo is constructed
Cell proliferation Cell specialization Cell interaction Cell movement

8
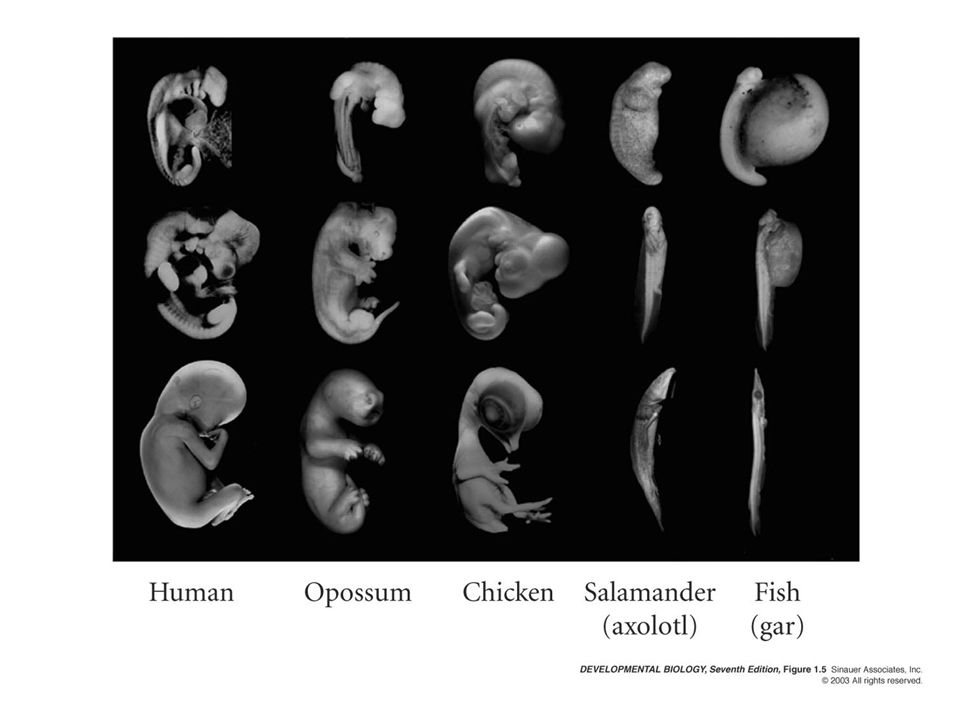
Similar basic anatomical features
Universal mechanisms of animal development Similar gene usage Similar basic anatomical features

9
Universal mechanisms of animal development-similar gene usage

10
Universal mechanisms of animal development-similar basic structure

12
Unicellular vs. Multicellular organisms
Tranmembrane proteins (e.g C. elegans genes over yeast) -Ion channels -Cell adhesion molecules -Cell surface receptors Gene regulatory proteins (e.g. HLH gene family: 141 humans, 84 fly, 41 C. elegans, 7 in yeast)
 -Ion channels. -Cell adhesion molecules. -Cell surface receptors. Gene regulatory proteins. (e.g. HLH gene family: 141 humans, 84 fly, 41 C. elegans, 7 in yeast)")
13
How species can be different?
Different animals utilize similar collection of genes Species identity genes (e. g 1% between human and chimpanchee ) non-coding, regulatory DNA sequences are highly differential between species
 non-coding, regulatory DNA sequences are highly differential between species.")
14
Different genome causes different behaviors of cells
Urchin Urchin Frog Fly, Mouse Frog 8.5

15
Cell fate and morphogenesis

16
Cell fate determination

17
Determination (결정) Differentiation (분화된 상태)
= formation of specialized cell types Commitment (예정된 상태) = biochemical changes in a cell that restrict its developmental fate
 = biochemical changes in a cell that restrict its developmental fate.")
18
Arms vs. Legs: Differential Gene Expression
Two transcription factors: Tbx5: Forelimbs Tbx4: Hindlimbs Expression dependant on anterior/posterior location Tbx4 in leg bud Tbx5 in wing bud

19
Cell fate commitment by genes

20
Tbx5 Tbx4

21
How cell fate is determined?
Autonomous specification Regulative specification

22
Commitment Three modes of initiating commitment have been described.
Autonomous Specification Conditional Specification Syncytial Specification

23
Autonomous Specification
= cell fate is determined before fertilization my morphogenetic determinants in ovum. Morphogenetic Determinants = mRNA or proteins that cause cellular commitment Mosaic Development = embryo functions like a “mosaic” of independent self-differentiating parts.

24
8.22 b-Catenin positive cells Asymmetric division

25
Mosaic Development determinant zygote

26
Mosaic Development determinant zygote Dies or 1/2 Embryo Forms

27
Conditional Specification
Conditional Specification = cell fate is determined by the conditions surrounding the cell. Morphogenetic determinants produced by cells within the embryo. (signaling among cells) Regulative Development = cells of an embryo can change fate based on the conditions within the embryo.
 Regulative Development = cells of an embryo can change fate based on the conditions within the embryo.")
28
Regulative Development
Normal Embryo Forms zygote

29
Regulative Development

30
Syncytial Specification
Syncytial Specification = cell fate is determined by the conditions affecting nuclei in a single multinucleate cell. Syncytium = a cytoplasm containing multiple nuclei. Morphogens may form a gradient within the cytoplasm.

31
Basic mechanisms of cell fate determination
Inductive interaction Morphogen Extracellular inhibitor Intrinsic program for time course Lateral inhibition

32
Basic mechanisms of cell fate determination
Inductive interaction Morphogen Extracellular inhibitor Intrinsic program for time course

33
Morphogen = Soluble molecule that causes cellular commitment but is secreted some distance from the target cells. Morphogen Gradient = concentration gradient of a morphogen.

34
Morphogen Threshold Concentrations
embryo Morpho- gen conc. position p. 63

35
Morphogen Threshold Concentrations
embryo Morpho- gen conc. position p. 63

36
Morphogen Threshold Concentrations
embryo Morpho- gen conc. position p. 63

37
Influence of Other Cells
Morphogen Receptor Gradient = frequency gradient of the receptors for a morphogen in target cell cell membranes. Morphogen gradient Morphogen receptor gradient

38
Activin Gradient Activin = morphogen in frog blastula, morphogen gradient of activin commits cells as a type of mesoderm. No activin = ectoderm heart cells noto- chord muscle blood

39
Frog Blastula (section)
ectoderm mesoderm endoderm blastocoel vegetal pole

40
Morphogenetic Field Morphogenetic Field = a group of cells whose position and fate are specified with respect to the same set of boundaries. Within a morphogenetic field fate is not yet specified. The limb field will form a limb. If divided the limb field will form two limbs.

41
Sonic Hedgehog

42
Ahn and Joyner (2004) Cell, 118, Gli1-CRE + LacZ conditional expression +/+ Gli2-/- Gli3-/-
 Cell, 118, Gli1-CRE + LacZ conditional expression +/+ Gli2-/- Gli3-/-")
43
Basic mechanisms of cell differentiation
Inductive interaction Morphogen Extracellular inhibitor Intrinsic program for time course

44
Basic mechanisms of cell differentiation
Inductive interaction Morphogen Extracellular inhibitor Intrinsic program for time course -Time keeping mechanisms -Cell division associated -Glial progenitor cells become oligodendrocytes after 8 divisions

45
Morphogenesis Commitment Cell shape changes. Cell movement.
Cell death. Changes in cell membranes or secreted products.

46
Sequential induction makes complex patterns

47
General Cell Types Epithelial Cells = cells connected together in sheets (attached to each other and an acellular basal lamina). fold, elevate, expand, involute, intercalate Mesenchymal Cells = cells unconnected together and operate independently. ingress, migrate

48
Epithelial Fold

49
Epithelial Fold

50
Epithelial Fold

51
Cell Affinity

52
Cell Affinity

53
Cell Affinity Selective Affinity = Disassociated cells will group together with (positive affinity) or will not group together with (negative affinity) only certain other cells. Homotropic Aggregation = Disassociated cells of the same type group together. (positive afinity)
 or will not group together with (negative affinity) only certain other cells. Homotropic Aggregation = Disassociated cells of the same type group together. (positive afinity)")
54
Cell Adhesion Differential Adhesion Hypothesis = explains patterns of cell sorting based on thermodynamics of affinity between adhesion molecules. Surface tension. Different adhesion molecules. Different amounts of the same type of adhesion molecules.

55
2 different amounts of the same adhesion molecule
Cell Affinity 2 different adhesion molecules 2 different amounts of the same adhesion molecule

56
Cell Adhesiveness Adhesion molecules = proteins in cell membrane.
Cadherins (5 classes) calcium dependent adhesion molecules binds to other cadherins (same type) connected to cytoskeleton by catenins Homophilic binding = adhesion molecules attach to the same class of adhesion molecule.
 calcium dependent adhesion molecules. binds to other cadherins (same type) connected to cytoskeleton by catenins. Homophilic binding = adhesion molecules attach to the same class of adhesion molecule.")
57
Cadherins cell membrane Ca2+ catenins actin (cytoskeleton) p. 72
 p. 72")
58
Cell Affinity N-cadherin E-cadherin

59
Methods for developmental biology
Descriptive embryology Experimental embryology Developmental genetics

60
Origins of Descriptive Embryology
Epigenesis vs. Preformationism preformationism argued for species continuity and constancy to some, epigenesis implied a need for a mysterious vital “life force” that was required to create life de novo careful observations on the anatomical development of embryos eventually required acceptance of epigenetic development

61
Classical Embryology Kaspar Wolff (1767): studies of chick embryogenesis Where did the instructions to build the embryo come from? Were they internal or external? ‘vital force’ [vis essentialis] needed to explain embryonic organization?

62
Classical Embryology Christian Pander (1774-1865)
studied the chick embryo and identified primary germ layers found in triploblastic embryos ectoderm: gives rise to outer layer of embryo and nervous system endoderm: gives rise to innermost layer and gives rise to digestive tube and associated organs mesoderm: middle layer that gives rise to bones, connective tissues, kidney, gonads, heart and hematopoietic system primary germ layers interact to form organs

63
Classical Embryology Karl Ernst Von Baer (1792-1896)
“enwicklungsgeshicte”: extended Pander’s observations; discovered notochord his work on chick embryogenesis was death knell to preformationism (also discovered mammalian egg)
")
64
Classical Embryology Von Baer’s laws:
the general features of a large group of animals appear earlier in development than specialized features in a small group within embryos, specialized structures develop from more generalized structures an embryo does not “pass through” the adult stages observed in lower animals: ontogeny does not recapitulate phylogeny early embryos share characteristics in common and become more and more divergent as development proceeds

65
Classical Embryology Wilhelm His (1831-1904)
one of the major antagonists to Haeckel developed the microtome, allowing for serial sectioning and much better anatomical resolution and reconstruction focused on the the mechanics of development and the importance of morphogenic movements, foldings and cellular interactions in the process of development.

66
The birth of experimental embryology
Defect = destroy part of embryo. Isolation = remove part of embryo and observe its development in culture. Recombination = replace part of an embryo with a part of the same embryo. Transplantation = replace part of an embryo with a part from a different embryo.

67
Birth of Experimental Embryology
Laurent Chabry (1887) ‘Qualitative mosaic’ experiments performed by isolating specific cells in developing tunicate embryos each blastomere was responsible for producing a particular set of larval tissues the blastomeres were apparently developing autonomously mosaic development: embryo constructed of individual modules capable of self-differentiation
 ‘Qualitative mosaic’ experiments performed by isolating specific cells in developing tunicate embryos. each blastomere was responsible for producing a particular set of larval tissues. the blastomeres were apparently developing autonomously. mosaic development: embryo constructed of individual modules capable of self-differentiation.")
68
Birth of Experimental Embryology
Wilhelm Roux ( ) ‘Quantitative mosaic’ student of Haeckel who performed ablation experiments in frogs Result of fate mapping in frogs implied that the destruction of certain regions in the early blastula would preclude development of certain structures destroyed right or left halves of frog embryos at 2 and 4 cell stages obtained “half embryos” having a complete right or left side, arguing for a mosaic model of development
 ‘Quantitative mosaic’ student of Haeckel who performed ablation experiments in frogs. Result of fate mapping in frogs implied that the destruction of certain regions in the early blastula would preclude development of certain structures. destroyed right or left halves of frog embryos at 2 and 4 cell stages. obtained half embryos having a complete right or left side, arguing for a mosaic model of development.")
69
Birth of Experimental Embryology
Hans Driesch ( ): ‘Regulative development’ each of the blastomeres from a two cell embryo developed into a complete larvae some of the later stage cells also developed into complete larvae conflicts with experiments of Roux and Chabry: first example of regulative development
: ‘Regulative development’ each of the blastomeres from a two cell embryo developed into a complete larvae. some of the later stage cells also developed into complete larvae. conflicts with experiments of Roux and Chabry: first example of regulative development.")
70
Birth of Experimental Embryology
Hans Driesch ( ): pressure plate experiment; by compressing the developing embryo between two plates, he could force a change in cleavage plane from equatorial to meridional, resulting in a different pattern of cleavage from normal. This reshuffled the position of the nuclei in the embryo…did it alter the fate map? Embryos were normal
: pressure plate experiment; by compressing the developing embryo between two plates, he could force a change in cleavage plane from equatorial to meridional, resulting in a different pattern of cleavage from normal. This reshuffled the position of the nuclei in the embryo…did it alter the fate map Embryos were normal.")
71
Birth of Experimental Embryology
Pressure plate experiments implied: nuclear equivalence cytoplasmic/nuclear interactions Driesch left science as a result of these experiments; he could not explain these results relative to the physics of his day and came to the philosophical view that living things can not be explained solely through physical laws

72
Experimental Design Matters!
J. F. McClendon (1910) Repeated experiments in frog development using Driesch’s isolation technique relative to Roux’s ablation technique noted regulative development NOT mosaic development isolated frog blastomeres developed into a whole frog ablated blastomeres were still in contact with intact blastomeres; they still were providing information for developmental programming
 Repeated experiments in frog development using Driesch’s isolation technique relative to Roux’s ablation technique. noted regulative development NOT mosaic development. isolated frog blastomeres developed into a whole frog. ablated blastomeres were still in contact with intact blastomeres; they still were providing information for developmental programming.")
73
Fate Mappng Fate maps do not necessarily imply commitment; not maps of potency or states of determination clonal restriction does not imply determination: allocation: clonal restriction in a population regardless of state of commitment commitment: intrinsic aspect of a cell that makes it follow a particular developmental path ‘commitment’ vs. ‘determination’?

74
The birth of developmental genetics
C. elegans, Drosophila, Frog, Mice, Plant as model systems

Similar presentations










 How do oogenesis and spermatogenesis differ? (Ch. 46) How do these hormones affect the menstrual cycle? LH FSH Estrogen Progesterone.>")








