Download presentation
Presentation is loading. Please wait.

1
Diseño de primers

2
Francis Crick Alex Rich Leslie Orgel James Watson

3
Escenarios frecuentes en el diseño de primers: Secuencia de DNA conocida –Diagnóstico, genotipificación –Tener cuidado si es DNA o cDNA Secuencia de DNA desconocida –Se busca secuenciar el gen en cuestión –Regiones conservadas en genes homólogos de organismos cercanos –Extremos amino ó carboxilo terminal de la proteína codificada es conocida

4
Secuencia de DNA conocida Los primers se diseñan a partir de criterios de optimalidad: –Estabilidad térmica del complejo primer-DNA –Estructura secundaria –Dímeros –Temperatura media de fusión –Especificidad

6
PCR Heat Cool Heat etc. 2 copies + and - strands of template forward primer reverse primer amplified DNA 3’ 5’ 3’ 5’ 3’ 5’ 4 copies 8 copies Heat Cool 1 copy

7
Ciclos deTemperatura Temperatura de hibridación (annealing) (usual 45-60 C) allows primers to hybridize to template Temperatura de extensión (usually 72 C) allows polymerase to extend starting at the primer Temperatura de denaturación (usually 95 C) separates strands
 (usual C) allows primers to hybridize to template Temperatura de extensión (usually 72 C) allows polymerase to extend starting at the primer Temperatura de denaturación (usually 95 C) separates strands")
8
Tiempo de vida media de Taq polymerasa aprox. 30 min a 95°C, Es lar razón por la que después de 30 ciclos de amplificación se pierde eficiencia Es posible reducir la temperatura de denaturación luego de 10 ciclos de amplificación ya que la longitud media del DNA blanco se ha reducido cercanamente al tamaño del amplicón: para ‘templados’ de 300bp o menos, la temperatura de denaturación puede reducirse a valores tan bajos como 88°C para un 50% de (G+C) (Yap and McGee, 1991), por lo que es posible llegar hasta 40 ciclos sin disminuir sustancialmente la eficiencia de la enzima.
 (Yap and McGee, 1991), por lo que es posible llegar hasta 40 ciclos sin disminuir sustancialmente la eficiencia de la enzima..")
9
Durante el PCR se da una liberación de pirofosfato que juntamente con el producto van a comportarse de manera inhibitoria

10
Considerations About Primers Specificity Specific for the intended target sequence (avoid nonspecific hybridization) Stability Form stable duplex with template under PCR conditions Compatibility Primers used as a pair shall work under the same PCR condition Uniqueness Length Annealing Temperature Primer Pair Matching Internal Structure Base Composition Internal Stability Characteristics of primers:Thoughts on primer design: Melting Temperature
 Stability Form stable duplex with template under PCR conditions Compatibility Primers used as a pair shall work under the same PCR condition Uniqueness Length Annealing Temperature Primer Pair Matching Internal Structure Base Composition Internal Stability Characteristics of primers:Thoughts on primer design: Melting Temperature")
11
Uniqueness There shall be one and only one target site in the template DNA where the primer binds, which means the primer sequence shall be unique in the template DNA. There shall be no annealing site in possible contaminant sources, such as human, rat, mouse, etc. (BLAST search against corresponding genome) Primer candidate 1 5’-TGCTAAGTTG-3’ Primer candidate 2 5’- CAGTCAACTGCTAC-3’ TGCTAAGTTG CAGTCAACTGCTAC Template DNA 5’...TCAACTTAGCATGATCGGGTA...GTAGCAGTTGACTGTACAACTCAGCAA...3’ NOT UNIQUE! UNIQUE! TGCT AGTTG A
 Primer candidate 1 5’-TGCTAAGTTG-3’ Primer candidate 2 5’- CAGTCAACTGCTAC-3’ TGCTAAGTTG CAGTCAACTGCTAC Template DNA 5’...TCAACTTAGCATGATCGGGTA...GTAGCAGTTGACTGTACAACTCAGCAA...3’ NOT UNIQUE. UNIQUE. TGCT AGTTG A.")
12
Length Primer length has effects on uniqueness and melting/annealing temperature. Roughly speaking, the longer the primer, the more chance that it’s unique; the longer the primer, the higher melting/annealing temperature. Generally speaking, the length of primer has to be at least 15 bases to ensure uniqueness. Usually, we pick primers of 17-28 bases long. This range varies based on if you can find unique primers with appropriate annealing temperature within this range.

13
Base Composition Base composition affects hybridization specificity, melting/annealing temperature and internal stability. Random base composition is preferred. We shall avoid long (A+T) and (G+C) rich region if possible. Usually, average (G+C) content around 50-60% will give us the right melting/annealing temperature for ordinary PCR reactions, and will give appropriate hybridization stability. However, melting/annealing temperature and hybridization stability are affected by other factors, which we’ll discuss later. Therefore, (G+C) content is allowed to change. Template DNA 5’...TCAACTTAGCATGATCGGGCA...AAGATGCACGGGCCTGTACACAA...3’ GCCCG TGCCCGATCATGCT GCCCG GCCCG CAT T T AT GC
 and (G+C) rich region if possible. Usually, average (G+C) content around 50-60% will give us the right melting/annealing temperature for ordinary PCR reactions, and will give appropriate hybridization stability. However, melting/annealing temperature and hybridization stability are affected by other factors, which we’ll discuss later. Therefore, (G+C) content is allowed to change. Template DNA 5’...TCAACTTAGCATGATCGGGCA...AAGATGCACGGGCCTGTACACAA...3’ GCCCG TGCCCGATCATGCT GCCCG GCCCG CAT T T AT GC.")
14
Thermodynamics Gibbs Free Energy, G Describes the energetics of biomolecules in aqueous solution. The change in free energy, G, for a chemical process, such as nucleic acid folding, can be used to determine the direction of the process: G=0: equilibrium G>0: unfavorable process G<0: favorable process Thus the natural tendency for biomolecules in solution is to minimize free energy of the entire system (biomolecules + solvent). G = H - T S H is enthalpy, S is entropy, and T is the temperature in Kelvin. Molecular interactions, such as hydrogen bonds, van der Waals and electrostatic interactions contribute to the H term. S describes the change of order of the system. Thus, both molecular interactions as well as the order of the system determine the direction of a chemical process. For any nucleic acid solution, it is extremely difficult to calculate the free energy from first principle Biophysical methods can be used to measure free energy changes
. G = H - T S H is enthalpy, S is entropy, and T is the temperature in Kelvin. Molecular interactions, such as hydrogen bonds, van der Waals and electrostatic interactions contribute to the H term. S describes the change of order of the system. Thus, both molecular interactions as well as the order of the system determine the direction of a chemical process. For any nucleic acid solution, it is extremely difficult to calculate the free energy from first principle Biophysical methods can be used to measure free energy changes.")
15
Internal Stability Stability Profile: Internal stability is calculated with entropy values of neighbor nucleotides. Usually, We draw a graph of ΔG for all nucleotides of the primers. This is known as the stability profile.

16
3’ Stability & 5’ Stability Template DNA 5’...TCAACTTAGCGTGATCGATTA...AAGATTCGCGTTAGCTGTACACAA...3’ 5’-AATCGATCTCGC-3’ CGC-3’ 5’-AATCGATCT This feature can be utilized for modifying 5’ end so that there will be a restriction enzyme site Primer elongation starts at 3’ end. Therefore, as long as 3’ end hybridizes to the template stably, the elongation begins. 5’ end sequence plays less important role. This feature can be utilized for modifying 5’ end so that there will be a restriction enzyme site. However, this feature also brings out a problem – if 3’end of the primer has 3 or more than 3 C/G, it can almost bind stably to any site where there are 3 complement G/C bases. Ideal situation will be stable 5’-termini + less stable 3’-termini, which eliminates false priming due to annealing of 3'-half of primer only. We prefer the 5’ end has 1 or two G/C bases (GC clamp) and the 3’ end has no more than 1 G/C base.
 and the 3’ end has no more than 1 G/C base..")
17
Melting Temperature Melting Temperature, Tm – the temperature at which half the DNA strands are single stranded and half are double-stranded.. Tm is characteristics of the DNA composition; Higher G+C content DNA has a higher Tm due to more H bonds. Calculation Method 1 (Base Composition): Tm = ( A + T ) 2 C + ( C + G ) 4 C (<20bp) Method 2 (Salt Adjusted): Used by GCG and Primer3 Tm = 81.5 + 16.6 [log 10 [Na+]] + 0.41 [GC%] – 0.65 [formamide%] – 675/length – mismatch% Method 3 (Nearest Neighbor): Used by OLIGO H / (10.8 + S + R ln (c / 4)) – 273.15 + 16.6(log 10 [K+]) where H is the sum of enthalpy of the nearest neighbors, S is the sum of entropy of the nearest neighbors, c is the molar concentration of primer, and R is the gas constant (1.987). As you can tell from the equation – higher G+C, higher Tm; Higher [probe], higher Tm; higher [K+], higher Tm Differences are sometimes significant, like 8 degrees, and sometimes trivial, like 0.1 degree. Try different software to calculate T m. Pick the common value.
: Tm = ( A + T ) 2 C + ( C + G ) 4 C (<20bp) Method 2 (Salt Adjusted): Used by GCG and Primer3 Tm = [log 10 [Na+]] [GC%] – 0.65 [formamide%] – 675/length – mismatch% Method 3 (Nearest Neighbor): Used by OLIGO H / ( S + R ln (c / 4)) – (log 10 [K+]) where H is the sum of enthalpy of the nearest neighbors, S is the sum of entropy of the nearest neighbors, c is the molar concentration of primer, and R is the gas constant (1.987). As you can tell from the equation – higher G+C, higher Tm; Higher [probe], higher Tm; higher [K+], higher Tm Differences are sometimes significant, like 8 degrees, and sometimes trivial, like 0.1 degree. Try different software to calculate T m. Pick the common value..")
18
Annealing Temperature T anneal = T m_primer – 4 C or 0.3 T m_primer + 0.7 T m_product – 14.9 Where T m_primer is the melting temperature for primer and T m_product is the melting temperature for product. Annealing Temperature, T anneal – the temperature at which primers anneal to the template DNA. It can be calculated from T m. To ensure that primers anneal to the template before the two strands of template anneal to each other, it’ required that the T m_product – T anneal ≥ 30 C

19
Stringency in Primer Annealing Stringency determines the specificity of the amplified DNA product. T anneal is the most significant factor affecting the stringency in primer annealing. T anneal : too low less stringent primer matches elsewhere too high more stringent primer may fail to match Other factors: GC%: GC pairs are more stringent than AT paris Salt & Buffer

20
Internal Structure If primers can anneal to themselves, or anneal to each other rather than anneal to the template, the PCR efficiency will be decreased dramatically. They shall be avoided. However, sometimes these 2 structures are harmless when the annealing temperature does not allow them to take form. For example, some dimers or hairpins form at 30 C while during PCR cycle, the lowest temperature only drops to 60 C.

21
Primer Pair Matching Primers work in pairs – forward primer and reverse primer. Since they are used in the same PCR reaction, it shall be ensured that the PCR condition is suitable for both of them. One critical feature is their annealing temperatures, which shall be compatible with each other. The maximum difference allowed is 3 C. The closer their T anneal are, the better.

22
Calculation procedure for extinction (absorption) coefficient of DNA Extinction coefficient at 260 nm, 25 degrees of Celsius, and neutral pH for the single-strand DNA is determined by the nearest-neighbor method
 coefficient of DNA Extinction coefficient at 260 nm, 25 degrees of Celsius, and neutral pH for the single-strand DNA is determined by the nearest-neighbor method")
23
The following table contains extinction coefficients [l/(mmol.cm)]: stack or monomerextinction coefficient pdA15.4 pdC7.4 pdG11.5 pdT8.7 dApdA13.7 dApdC10.6 dApdG12.5 dApdT11.4 dCpdA10.6 dCpdC7.3 dCpdG9.0 dCpdT7.6 dGpdA12.6 dGpdC8.8 dGpdG10.8 dGpdT10.0 dTpdA11.7 dTpdC8.1 dTpdG9.5 dTpdT8.4
![The following table contains extinction coefficients [l/(mmol.cm)]: stack or monomerextinction coefficient pdA15.4 pdC7.4 pdG11.5 pdT8.7 dApdA13.7 dApdC10.6 dApdG12.5 dApdT11.4 dCpdA10.6 dCpdC7.3 dCpdG9.0 dCpdT7.6 dGpdA12.6 dGpdC8.8 dGpdG10.8 dGpdT10.0 dTpdA11.7 dTpdC8.1 dTpdG9.5 dTpdT8.4](http://images.slideplayer.com/14/4299279/slides/slide_23.jpg "The following table contains extinction coefficients [l/(mmol.cm)]: stack or monomerextinction coefficient pdA15.4 pdC7.4 pdG11.5 pdT8.7 dApdA13.7 dApdC10.6 dApdG12.5 dApdT11.4 dCpdA10.6 dCpdC7.3 dCpdG9.0 dCpdT7.6 dGpdA12.6 dGpdC8.8 dGpdG10.8 dGpdT10.0 dTpdA11.7 dTpdC8.1 dTpdG9.5 dTpdT8.4")
24
Summary ~ when is a “primer” a primer? 5’3’ 5’ 3’

25
Summary ~ Primer Design Criteria 1.Uniqueness: ensure correct priming site; 2.Length: 17-28 bases.This range varies; 3.Base composition: average (G+C) content around 50-60%; avoid long (A+T) and (G+C) rich region if possible; 4.Optimize base pairing: it’s critical that the stability at 5’ end be high and the stability at 3’ end be relatively low to minimize false priming. 5.Melting Tms between 55-80 C are preferred; 6.Assure that primers at a set have annealing Tm within 2 – 3 C of each other. 7.Minimize internal secondary structure: hairpins and dimmers shall be avoided.

26
Computer-Aided Primer Design artfar better done by machines Primer design is an art when done by human beings, and a far better done by machines. Some primer design programs we use: - Oligo: Life Science Software, standalone application - GCG: Accelrys, ICBR maintains the server. - Primer3: MIT, standalone / web application http://www-genome.wi.mit.edu/cgi-bin/primer/primer3_www.cgi - BioTools: BioTools, Inc. ICBR distributes the license. - Others: GeneFisher, Primer!, Web Primer, NBI oligo program, etc. Melting temperature calculation software: - BioMath: http://www.promega.com/biomath/calc11.htmhttp://www.promega.com/biomath/calc11.htm

27
Primer Design on the Web There are a bunch of good PCR primer design programs on the web: –Primer 3 at the MIT Whitehead Institute http://www.genome.wi.mit.edu/cgi-bin/primer/primer3_www.cgi –Cassandra at the Univ. of Southern California http://www- hto.usc.edu/software/procrustes/cassandra/cass_frm.html –GeneFisher by Folker Meyer & Chris Schleiermacher at Bielefeld University, Germany http://bibiserv.TechFak.Uni-Bielefeld.DE/genefisher/ –Xprimer at the Virtual Genome Center, Univ. Minnesota Medical School http://alces.med.umn.edu/rawprimer.html

28
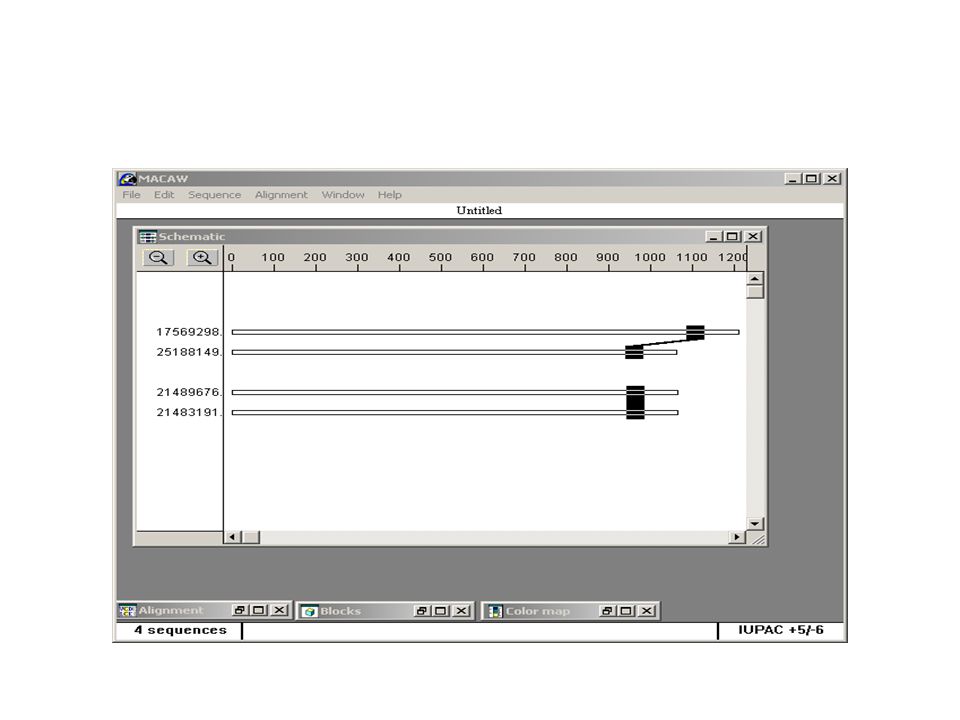
Secuencia de DNA desconocida Regiones conservadas en genes homólogos de organismos cercanos

29
>gi|17563797:1-1014 Caenorhabditis elegans essential CathePsin L (38.1 kD) (cpl-1), mRNA ATGAACCGATTCATTCTTCTGGCACTGGTTGCCGCCGTCGTCGCCGTCAATTCGGCCAAGCTGTCCCGTC AAATCGAGTCGGCCATCGAGAAATGGGACGACTATAAGGAGGACTTTGATAAGGAGTACTCGGAGAGCGA GGAGCAGACCTATATGGAGGCATTTGTCAAGAATATGATCCATATTGAGAATCACAACAGAGATCACCGA CTCGGAAGAAAGACATTCGAGATGGGATTGAATCATATTGCTGACTTGCCATTCAGCCAATACCGCAAAC TCAACGGTTACAGACGTCTCTTCGGTGACTCCAGAATCAAGAACTCCTCCTCTTTCTTGGCTCCATTCAA TGTTCAGGTCCCAGATGAGGTTGACTGGCGTGATACCCACCTCGTCACTGATGTCAAGAACCAAGGAATG TGCGGATCGTGCTGGGCCTTCTCCGCCACCGGAGCCCTCGAAGGACAACACGCTCGCAAGCTGGGACAAC TCGTCTCCCTTTCCGAGCAAAACCTCGTCGACTGCTCTACCAAGTACGGAAACCACGGATGCAACGGAGG ACTCATGGATCAAGCTTTCGAGTACATTCGTGACAACCATGGTGTCGACACCGAGGAGTCATACCCATAC AAGGGACGTGACATGAAGTGTCACTTCAACAAGAAGACCGTCGGAGCTGATGATAAGGGATACGTTGACA CCCCAGAAGGAGATGAGGAGCAACTTAAGATCGCTGTCGCCACCCAAGGACCAATCTCTATTGCTATCGA CGCCGGACACCGCAGCTTCCAACTTTACAAGAAGGGAGTCTACTACGATGAGGAATGCTCATCCGAAGAG CTCGACCACGGAGTGCTTCTCGTCGGATACGGAACCGACCCAGAGCACGGAGACTACTGGATTGTCAAGA ACTCGTGGGGAGCTGGATGGGGAGAGAAGGGATACATCCGTATCGCCCGTAACCGCAACAATCACTGCGG AGTCGCCACCAAGGCCAGTTATCCATTGGTCTAA
 (cpl-1), mRNA ATGAACCGATTCATTCTTCTGGCACTGGTTGCCGCCGTCGTCGCCGTCAATTCGGCCAAGCTGTCCCGTC AAATCGAGTCGGCCATCGAGAAATGGGACGACTATAAGGAGGACTTTGATAAGGAGTACTCGGAGAGCGA GGAGCAGACCTATATGGAGGCATTTGTCAAGAATATGATCCATATTGAGAATCACAACAGAGATCACCGA CTCGGAAGAAAGACATTCGAGATGGGATTGAATCATATTGCTGACTTGCCATTCAGCCAATACCGCAAAC TCAACGGTTACAGACGTCTCTTCGGTGACTCCAGAATCAAGAACTCCTCCTCTTTCTTGGCTCCATTCAA TGTTCAGGTCCCAGATGAGGTTGACTGGCGTGATACCCACCTCGTCACTGATGTCAAGAACCAAGGAATG TGCGGATCGTGCTGGGCCTTCTCCGCCACCGGAGCCCTCGAAGGACAACACGCTCGCAAGCTGGGACAAC TCGTCTCCCTTTCCGAGCAAAACCTCGTCGACTGCTCTACCAAGTACGGAAACCACGGATGCAACGGAGG ACTCATGGATCAAGCTTTCGAGTACATTCGTGACAACCATGGTGTCGACACCGAGGAGTCATACCCATAC AAGGGACGTGACATGAAGTGTCACTTCAACAAGAAGACCGTCGGAGCTGATGATAAGGGATACGTTGACA CCCCAGAAGGAGATGAGGAGCAACTTAAGATCGCTGTCGCCACCCAAGGACCAATCTCTATTGCTATCGA CGCCGGACACCGCAGCTTCCAACTTTACAAGAAGGGAGTCTACTACGATGAGGAATGCTCATCCGAAGAG CTCGACCACGGAGTGCTTCTCGTCGGATACGGAACCGACCCAGAGCACGGAGACTACTGGATTGTCAAGA ACTCGTGGGGAGCTGGATGGGGAGAGAAGGGATACATCCGTATCGCCCGTAACCGCAACAATCACTGCGG AGTCGCCACCAAGGCCAGTTATCCATTGGTCTAA")
30
>gi|25188149:70-1131 Brugia malayi mRNA for cathepsin L-like cysteine proteinase, complete cds ATGAAGGCGTTTTTTATTCTACATTTAGCCAGTTTTCTCTTGCTTACTTATGCAAATCCACTTAATGAAC TGGATAATGATGACACACCAGGTAAAATTCATACCTTGTATCAGCAGCATTATTCGAAGTATAAAACATA TCTGAAAAAAATGGGCAAAAAACATGATCCATCTGTTCCGGAACCAATTCGATTACTTAAATTTGTACAA TCTTTGAAAATGATTGATGAACATAACCAGCGTTACAGTAAAGGATTGGAAACATACAAAGTAGATCTGA ACAAAATGAGCGATTGGACCGAAGAAGAGAAAGAAAGACTTCGGGGATATTATCCAAATTTGACTGAATA TGCGGAAGGAGATTTAAGTAGAATAATCCGAGGAAATATAACAACAACAATACCAAAGTCTTTTGATTAT AGAAAGAAAATAACTGTACTACCAGCATCAGATCAAGGTCGTTGTGGTGTGTGTTTTATTTTCTCAGCAT TAGGAGCGCTTGAAATGTATGTGGCGCTCAGGACTAAAAAACGGCCGGTAAAACTATCAGTACAGGATGT AATGGATTGTAGTGGAATGGAAAAATGCAAAGGAAGAGGCGGAAATGAACCTGCGGTTTTTCGTTGGGTT GCTGAGCACGGTGTAAAGACCGATAAGAGTTACCCATATAAGGAAAATGATAGTGTTTCATGCCCGAGAA ATACCCCACAACGACGAAAGTATGGTTTAGCTGATGCATTCTATTTACCTCCTAGCAATGAACAAATTCT CAAGAAGATACTAGCACTATATGGACCAGTTTGTGTATCGTTACATTCGTCATTACAAAGTTTTGTAGCT TATCGAAGTGGTATTTATAATGATCCAAAATGTCCAACTAATGCAGAAAAAGTAAATCATGCGGTAATAG CGGTTGGTTATGGTGTCCAGAATGGGATGGAATATTTTATCATCAAAAACAGTTGGGGCCCTACATGGGG TCAAAAAGGATACGGTCGTATTCGCGCTGGGGTATTTATGTGCGGTATTGGTCGTTTTTCGAACGTACCA ATCTTCAAATGA

31
>gi|21489676:4-1068 Haemonchus contortus cathepsin L cysteine protease (cpl-1) mRNA, complete cds ATGCTACGTCTGTTGTCGTTGGCGCTTCTATGCGCTGTAGTTTTAGCTAGTATTGATGGGTTCAGAAGGC ATGATCATGGCGTACGAGTGCACAGACAGAAAAGCCTTCGCCAAAAAATCGACGAGGCTTTCAATAAATG GGATGACTACAAGGAGACCTTTGGAAAGTCGTATGAACCGGATGAAGAGAACGACTACATGGAGGCCTTT GTGAAGAACGTGATTCACATTGAGGAACACAATAAGGAACACCGTCTTGGTAGGAAAACATTTGAAATGG GTCTCAACGAAATTGCTGATTTGCCATTCTCACAATATCGAAAACTCAACGGGTATCGTATGCGTCGTCA ATTTGGCGATTCCTTGCAGTCCAATGGTACCAAGTTTTTGGTTCCATTCAATGTTCAGATCCCGGAATCT GTTGACTGGCGAGAGGAAGGACTTGTGACTCCAGTAAAGAATCAAGGAATGTGTGGATCATGTTGGGCGT TCTCCTCTACTGGTGCTCTAGAAGGACAACATGCACGTGCCACTGGCAAGCTGGTATCCCTTTCCGAGCA AAATCTTGTCGATTGCTCAACGAAGTACGGAAACCATGGCTGCAACGGTGGTCTTATGGATTTGGCATTT GAATATATCAAGGAAAATCACGGTGTCGATACCGAAGATAGTTATCCATACGTCGGAAGAGAAACGAAAT GTCATTTCAAGAGGAACGCTGTTGGAGCCGATGACAAGGGCTTTGTAGATCTTCCTGAAGGCGATGAAGA GGCGTTGAAGAAAGCAGTTGCCACTCAAGGTCCAATTTCTATCGCTATTGACGCTGGTCACAGGTCATTC CAGCTGTACAAGAAGGGAGTGTACTTTGACGAAGAGTGCTCGTCTGAAGAACTGGATCACGGTGTTCTTC TCGTTGGATACGGTACTGATCCCGAGGCAGGAGATTATTGGCTTGTGAAGAACAGCTGGGGACCGACTTG GGGAGAGAAGGGATACATTCGCATTGCCCGTAACCGCAACAATCACTGCGGTGTTGCAACAAAGGCCAGC TACCCGCTCGTTTAG
 mRNA, complete cds ATGCTACGTCTGTTGTCGTTGGCGCTTCTATGCGCTGTAGTTTTAGCTAGTATTGATGGGTTCAGAAGGC ATGATCATGGCGTACGAGTGCACAGACAGAAAAGCCTTCGCCAAAAAATCGACGAGGCTTTCAATAAATG GGATGACTACAAGGAGACCTTTGGAAAGTCGTATGAACCGGATGAAGAGAACGACTACATGGAGGCCTTT GTGAAGAACGTGATTCACATTGAGGAACACAATAAGGAACACCGTCTTGGTAGGAAAACATTTGAAATGG GTCTCAACGAAATTGCTGATTTGCCATTCTCACAATATCGAAAACTCAACGGGTATCGTATGCGTCGTCA ATTTGGCGATTCCTTGCAGTCCAATGGTACCAAGTTTTTGGTTCCATTCAATGTTCAGATCCCGGAATCT GTTGACTGGCGAGAGGAAGGACTTGTGACTCCAGTAAAGAATCAAGGAATGTGTGGATCATGTTGGGCGT TCTCCTCTACTGGTGCTCTAGAAGGACAACATGCACGTGCCACTGGCAAGCTGGTATCCCTTTCCGAGCA AAATCTTGTCGATTGCTCAACGAAGTACGGAAACCATGGCTGCAACGGTGGTCTTATGGATTTGGCATTT GAATATATCAAGGAAAATCACGGTGTCGATACCGAAGATAGTTATCCATACGTCGGAAGAGAAACGAAAT GTCATTTCAAGAGGAACGCTGTTGGAGCCGATGACAAGGGCTTTGTAGATCTTCCTGAAGGCGATGAAGA GGCGTTGAAGAAAGCAGTTGCCACTCAAGGTCCAATTTCTATCGCTATTGACGCTGGTCACAGGTCATTC CAGCTGTACAAGAAGGGAGTGTACTTTGACGAAGAGTGCTCGTCTGAAGAACTGGATCACGGTGTTCTTC TCGTTGGATACGGTACTGATCCCGAGGCAGGAGATTATTGGCTTGTGAAGAACAGCTGGGGACCGACTTG GGGAGAGAAGGGATACATTCGCATTGCCCGTAACCGCAACAATCACTGCGGTGTTGCAACAAAGGCCAGC TACCCGCTCGTTTAG")
32
>(gi|21483189:<1-53, 1121-1209, 1586-1725, 1806-1910, 2049-2183, 2239-2385, 2448-2537, 2616-2691, 2999- 3107, 3188-3287) Dictyocaulus viviparus cathepsin L (cpl-1) gene, partial cds TTGCTTCTGTTCCTATGCGATTTAGCTTCAACTAAGATCGGTACACCTAGAAAGCACGGATTTTATTCGG AGAAACAGAAAAGTCTACGTCAGAGGATCGATGAGGCCTTTGGAAAATGGGATGAGTATAAGATCAAATA TGATAAACACTATGACCCTGAAGAAGAAAATGATTATATGGAGGCTTTTGTAAAAAACATGATCCACATC GAGGAGCATAATCATGAACACCGCTTGGGGCGAAAAACTTTCGAAATGGGATTGAACAATATCGCTGATC TTCCTTTCTCGGAGTACCGCAAATTGAACGGTTACCGCCATCGTCGCTTATTTGGTGACTCTATGCGTAA AAATGGCACGAAATTTTTAGTCCCTTTCAATGTCAAGGCGCCGGATTCAGTAGATTGGCGAGAACATAAT CTCGTTACTCCAGTAAAAAATCAAGGGATGTGTGGTTCCTGTTGGGCCTTCTCCGCAACTGGAGCTCTCG AAGGACAACATTTTCGTGCAACCGGAAAACTTGTTTCCTTGTCCGAACAGAATCTGGTGGATTGTTCTAC TAAATATGGAAACCATGGTTGTAACGGTGGTCTCATGGATTTGGCATTTGAATACATAAAAGATAATCAT GGTATAGATACTGAGGAGGGTTACCCTTATGTTGGCAAAGAGATGAGGTGCCACTTCAAAAAGAGGGACA TTGGAGCTGAAGATAGGGGGTTTGTAGATCTTCCAGAAGGAGACGAAGATGCCTTGAAGGTCGCTGTCGC TACTCAGGGTCCTATTTCTATTGCCATTGATGCCGGTCATCGGTCTTTCCAGTTATATAAAAAAGGAGTT TATTTTGACGAGGAGTGTTCATCCGAAGAACTTGATCATGGAGTTCTTCTTGTGGGTTATGGCACTGATC CTGAAGCTGGCGACTACTGGATCATAAAGAATAGTTGGGGAACTAAATGGGGAGAAAAGGGTTACGTTCG TATCGCTCGGAATCGCAACAATCACTGCGGTGTGGCAACGAAGGCGAGTTATCCGCTCGTCTAA
 Dictyocaulus viviparus cathepsin L (cpl-1) gene, partial cds TTGCTTCTGTTCCTATGCGATTTAGCTTCAACTAAGATCGGTACACCTAGAAAGCACGGATTTTATTCGG AGAAACAGAAAAGTCTACGTCAGAGGATCGATGAGGCCTTTGGAAAATGGGATGAGTATAAGATCAAATA TGATAAACACTATGACCCTGAAGAAGAAAATGATTATATGGAGGCTTTTGTAAAAAACATGATCCACATC GAGGAGCATAATCATGAACACCGCTTGGGGCGAAAAACTTTCGAAATGGGATTGAACAATATCGCTGATC TTCCTTTCTCGGAGTACCGCAAATTGAACGGTTACCGCCATCGTCGCTTATTTGGTGACTCTATGCGTAA AAATGGCACGAAATTTTTAGTCCCTTTCAATGTCAAGGCGCCGGATTCAGTAGATTGGCGAGAACATAAT CTCGTTACTCCAGTAAAAAATCAAGGGATGTGTGGTTCCTGTTGGGCCTTCTCCGCAACTGGAGCTCTCG AAGGACAACATTTTCGTGCAACCGGAAAACTTGTTTCCTTGTCCGAACAGAATCTGGTGGATTGTTCTAC TAAATATGGAAACCATGGTTGTAACGGTGGTCTCATGGATTTGGCATTTGAATACATAAAAGATAATCAT GGTATAGATACTGAGGAGGGTTACCCTTATGTTGGCAAAGAGATGAGGTGCCACTTCAAAAAGAGGGACA TTGGAGCTGAAGATAGGGGGTTTGTAGATCTTCCAGAAGGAGACGAAGATGCCTTGAAGGTCGCTGTCGC TACTCAGGGTCCTATTTCTATTGCCATTGATGCCGGTCATCGGTCTTTCCAGTTATATAAAAAAGGAGTT TATTTTGACGAGGAGTGTTCATCCGAAGAACTTGATCATGGAGTTCTTCTTGTGGGTTATGGCACTGATC CTGAAGCTGGCGACTACTGGATCATAAAGAATAGTTGGGGAACTAAATGGGGAGAAAAGGGTTACGTTCG TATCGCTCGGAATCGCAACAATCACTGCGGTGTGGCAACGAAGGCGAGTTATCCGCTCGTCTAA")
34
Universal Primers Primers can be designed to amplify only one product. Primers can also be designed to amplify multiple products. We call such primers “universal primers”. For example, design primers to amplify all HPV genes. Strategy: 1.Align groups of sequences you want to amplify. 2.Find the most conservative regions at 5’ end and at 3’ end. 3.Design forward primer at the 5’ conservative region. 4.Design reverse primer at the 3’ conservative regions. 5.Matching forward and reverse primers to find the best pair. 6.Ensure uniqueness in all template sequences. 7.Ensure uniqueness in possible contaminant sources.

40
Nucleic-acid Base Codes SymbolMeaningSymbolMeaning AASG or C GGWA or T CCHA, C, or T (~G) TTBC, G, or T (~A) RA or GVA, C, or G (~T) YC or TDA, G, or T (~C) MA or CNA, C, G, or T KG or T Adapted from Mount, Bioinformatics Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (2001)
 TTBC, G, or T (~A) RA or GVA, C, or G (~T) YC or TDA, G, or T (~C) MA or CNA, C, G, or T KG or T Adapted from Mount, Bioinformatics Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (2001)")
41
Catep1w 5'KTG YGG WKY VTG YTK KRY BTT CTC3' Catep2w 5'GTA WCC YTT YTS DCC CCA 3' Estos primers son las secuencias degeneradas de alinear los nematodes disponibles C.elegans, Brugian, Haemonchus, Dyctiocaulus

42
Secuencia de DNA desconocida –Extremos amino ó carboxilo terminal de la proteína codificada es conocida

43
P F T K CCn TTy ACn AAr Donde n = A,C,G or T y = C,T r = A,G

44
1 fold sitesM W 2 fold sitesF Y H Q N K D E C 3 fold sitesI 4 fold sitesV P T A G 6 fold sitesL S R

46
Our Focus: the Helix-Coil Transition in DNA In particular, we focus on two related processes: –DNA melting B helix to two coils. –DNA annealing two coils to a B helix. Understanding these: –aids in modeling more complicated transitions. e.g., many species.

47
Stabilizing Interactions DNA B-Helix structure stabilized by: –hydrogen bonding between bases (minor). –stacking between H + -bonded base-pairs (primary). induced dipole moments in the clouds of adjacent heterocyclic rings. stacking also sequesters hydrophobic rings. results in the characteristic helix shape. In DNA melting…the helix destabilized –generally implemented by increasing temperature, T. –destabilizes the stacks…unwinding the helix. –unwound helix separates into free ssDNAs (‘coils’).
. induced dipole moments in the clouds of adjacent heterocyclic rings. stacking also sequesters hydrophobic rings. results in the characteristic helix shape. In DNA melting…the helix destabilized –generally implemented by increasing temperature, T. –destabilizes the stacks…unwinding the helix. –unwound helix separates into free ssDNAs (‘coils’)..")
48
Monitoring the Helix-Coil Transition Degree of stacking experimentally observable: –Let B = mean fraction of stacked base pairs. –Ultraviolet absorbance at 260 nm (A 260 ) inversely proportional to B. the ‘hypochromicity’. –DNA melting accompanied by @ 40% increase A 260. –A 260 vs. T yields B vs. T (melting curve).
 inversely proportional to B. the ‘hypochromicity’. –DNA melting accompanied 40% increase A 260. –A 260 vs. T yields B vs. T (melting curve)..")
49
DNA Melting Curves B decreases monotonically from 1 to 0. –sigmoidal shape indicates DNA melting is cooperative. –Temp. at which B = ½ is the Melting temperature (T m ) –Width ( T) is non-zero (e.g., for 10-mers, T @ 10 o C). Melting curves of longer DNAs show more structure: –several independently melting regions (AT ’ s less stable). –melting curve then a combination of several sigmoids.
 –Width ( T) is non-zero (e.g., for 10-mers, 10 o C). Melting curves of longer DNAs show more structure: –several independently melting regions (AT ’ s less stable). –melting curve then a combination of several sigmoids..")
50
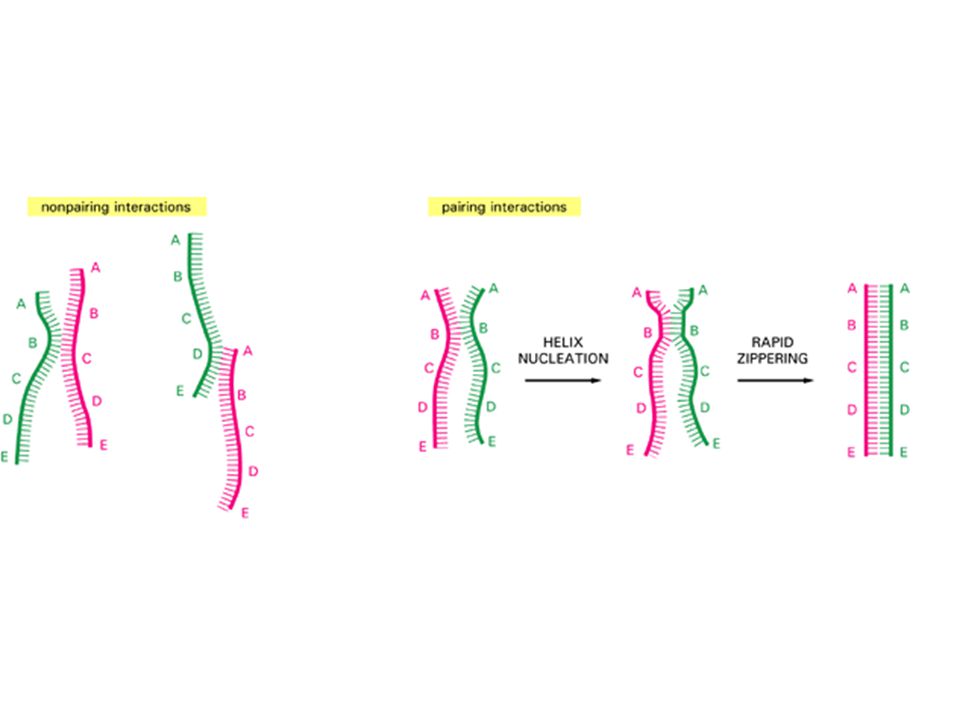
DNA Renaturation Renaturation is the ‘reverse’ of DNA melting. –also called DNA ‘annealing’ or ‘hybridization’. DNA renaturation is a much more complicated process:

51
DNA reassociation (renaturation) Double-stranded DNA Denatured, single-stranded DNA Slower, rate-limiting, second-order process of finding complementary sequences to nucleate base-pairing k2k2 Faster, zippering reaction to form long molecules of double- stranded DNA http://www3.kumc.edu/jcalvet/PowerPoint/bioc801b.ppt
 Double-stranded DNA Denatured, single-stranded DNA Slower, rate-limiting, second-order process of finding complementary sequences to nucleate base-pairing k2k2 Faster, zippering reaction to form long molecules of double- stranded DNA")
53
Reversibility of DNA Melting Melting for short DNAs strictly reversible. Reversibility of DNA melting: –measured by a lack of ‘hysteresis’ in the melting curve. –DNA melting curve = DNA renaturation curve. Validity of an equilibrium model of melting assumes: –melting slow enough to maintain equilibrium at each T. relatively slow heating/cooling (0.1-0.2 o C/minute). –failure to maintain equilibrium = hysteresis in the melting curve.
. –failure to maintain equilibrium = hysteresis in the melting curve..")
Similar presentations










 Calculating Melting Temperature (Tm) PCR Primers Design.>")


>")


 Presented by: Afsaneh Bazgir Polymerase Chain Reaction>")
 Lecture: PCR & primer stock preparation In-Class Writing: abstract for AEM 63: 2647-53, 1997 (page 68) Hand In: abstract.>")





>")


