Download presentation
Presentation is loading. Please wait.

1
C ONNECTIVE TISSUE Dr. Makarchuk Irina

2
C ONNECTIVE TISSUE COMPRISES A DIVERSE GROUP OF CELLS WITHIN A TISSUE - SPECIFIC EXTRACELLULAR MATRIX In general, connective tissue consists of cells and an extracellular matrix (ECM). ECM includes structural (fibers) and specialized proteins that constitute the ground substance. Connective tissue forms a vast and continuous compartment throughout the body, bounded by the basal laminae of the various epithelia and by the basal or external laminae of muscle cells and nerve- supporting cells.
 and specialized proteins that constitute the ground substance. Connective tissue forms a vast and continuous compartment throughout the body, bounded by the basal laminae of the various epithelia and by the basal or external laminae of muscle cells and nerve- supporting cells..")
3
C LASSIFICATION OF CONNECTIVE TISSUE IS BASED ON THE COMPOSITION AND ORGANIZATION OF ITS CELLULAR AND EXTRACELLULAR COMPONENTS AND ON ITS FUNCTIONS. Classification of Connective Tissue Embryonic connective tissue MesenchymeMucous connective tissue Connective tissue proper Loose connective tissueDense connective tissue Regular Irregular Specialized connective tissue Blood Hemopoietic tissue Adipose tissue Lymphatic tissue Cartilage Bone

4
B LOOD IS A FLUID CONNECTIVE TISSUE THAT CIRCULATES THROUGH THE CARDIOVASCULAR SYSTEM. Like the other connective tissues, blood consists of cells and an extracellular component. Total blood volume in the average adult is about 6 L or 7% to 8% of total body weight. The heart’s pumping action propels blood through the cardiovascular system to the body tissues. Blood’s many functions include: delivery of nutrients and oxygen directly or indirectly to cells, transport of wastes and carbon dioxide away from cells, delivery of hormones and other regulatory substances to and from cells and tissues, maintenance of homeostasis by acting as a buffer and participating in coagulation and thermoregulation, and transport of humoral agents and cells of the immune system that protect the body from pathogenic agents, foreign proteins, and transformed cells (i.e., cancer cells).
..")
5
B LOOD CONSISTS OF CELLS AND THEIR DERIVATIVES AND A PROTEIN RICH FLUID CALLED PLASMA. Blood cells and their derivatives include: erythrocytes, also called red blood cells (RBCs), leukocytes, also known as white blood cells (WBCs), and thrombocytes, also termed platelets.
, leukocytes, also known as white blood cells (WBCs), and thrombocytes, also termed platelets..")
6
P LASMA IS THE LIQUID EXTRACELLULAR MATERIAL THAT IMPARTS FLUID PROPERTIES TO BLOOD. The relative volume of cells and plasma in whole blood is approximately 45% and 55%, respectively. The volume of packed erythrocytes in a sample of blood is called the hematocrit. N.B. ! The hematocrit is measured by centrifuging a blood sample to which anticoagulants have been added, and then calculating the percentage of the centrifuge tube volume occupied by the erythrocytes compared with that of the whole blood. A normal hematocrit reading is about 39% to 50% in men and 35% to 45% in women; thus, 39% to 50% and 35% to 45% of the blood volume for men and women, respectively, consists of erythrocytes. Low hematocrit values often reflect reduced numbers of circulating erythrocytes (a condition called anemia) and may indicate significant blood loss caused by internal or external bleeding.
 and may indicate significant blood loss caused by internal or external bleeding..")
7
C OMPOSITION OF BLOOD PLASMA

8
F ORMED ELEMENTS OF THE BLOOD

9
ERYTHROCYTES Erythrocytes or red blood cells (RBCs), are anucleate cells devoid of typical organelles. They function only within the bloodstream to bind oxygen for delivery to the tissues and, in exchange, bind carbon dioxide for removal from the tissues. Their shape is that of a biconcave discs with a diameter of 7.8 µm, an edge thickness of 2.6 µm, and a central thickness of 0.8 µm. This shape maximizes the cell’s surface area (140 µm2), an important attribute in gas exchange. N.B.! The life span of erythrocytes is approximately 120 days, after which most (90%) of them are phagocytosed by macrophages in the spleen, bone marrow, and liver. The remaining aged erythrocytes (10%) break down intravascularly, releasing insignificant amounts of hemoglobin into the blood. Erythrocytes contain hemoglobin, a protein specialized for the transport of oxygen and carbon dioxide.
, an important attribute in gas exchange. N.B.. The life span of erythrocytes is approximately 120 days, after which most (90%) of them are phagocytosed by macrophages in the spleen, bone marrow, and liver. The remaining aged erythrocytes (10%) break down intravascularly, releasing insignificant amounts of hemoglobin into the blood. Erythrocytes contain hemoglobin, a protein specialized for the transport of oxygen and carbon dioxide..")
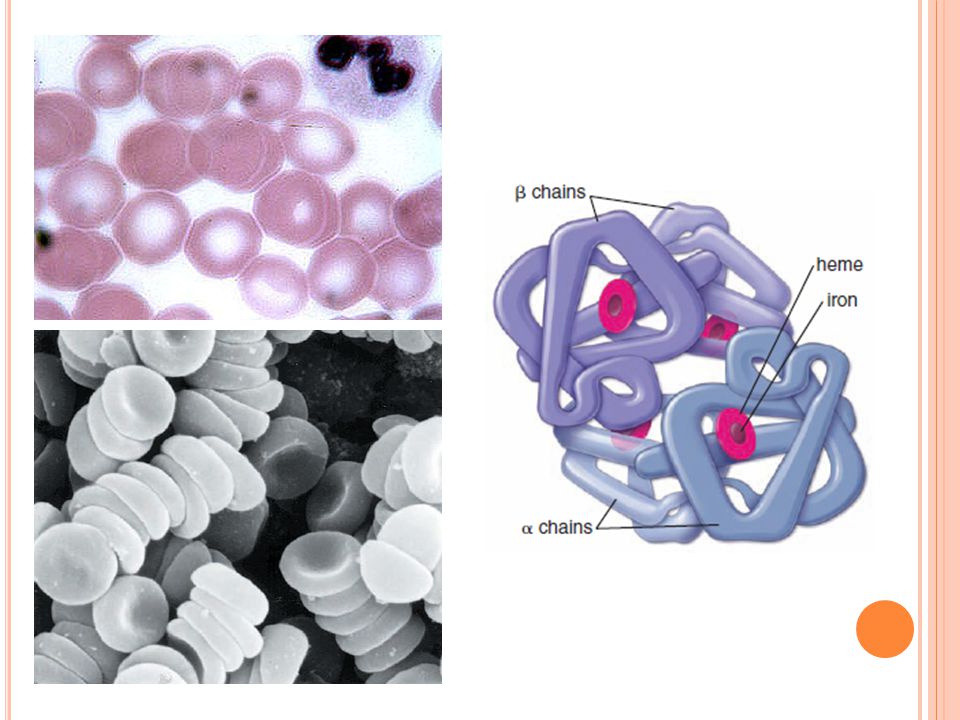
11
H EMOGLOBIN consists of four polypeptide chains of globin α, β, δ and γ, each complexed to an iron-containing heme group. The structure of the polypeptide chains varies. Depending on the particular polypeptides present, the following types of hemoglobin can be distinguished: Hemoglobin HbA is most prevalent in adults, accounting for about 96% of total hemoglobin. It is a tetramer with two α and two β chains (α2β2). Hemoglobin HbA2 accounts for 1.5% to 3% of total hemoglobin in adults. It consists of two α and two δ chains (α2δ2). Hemoglobin HbF accounts for less than 1% of total hemoglobin in adults. It contains two α and two δ chains (α2δ2) and is the principal form of hemoglobin in the fetus. N.B.! HbF production falls dramatically after birth; however, in some individuals HbF is produced throughout their entire lives. Although HbF persists in slightly higher percentages than normal in those with sickle cell disease and thalassemia, it does not appear to have a pathologic role.
. Hemoglobin HbA2 accounts for 1.5% to 3% of total hemoglobin in adults. It consists of two α and two δ chains (α2δ2). Hemoglobin HbF accounts for less than 1% of total hemoglobin in adults. It contains two α and two δ chains (α2δ2) and is the principal form of hemoglobin in the fetus. N.B.. HbF production falls dramatically after birth; however, in some individuals HbF is produced throughout their entire lives. Although HbF persists in slightly higher percentages than normal in those with sickle cell disease and thalassemia, it does not appear to have a pathologic role..")
12
LEUKOCYTES Leukocytes are subclassified into two general groups. The basis for this division is the presence or absence of prominent specific granules in the cytoplasm. As previously noted, cells containing specific granules are classified as granulocytes (neutrophils, eosinophils, and basophils), and cells that lack specific granules are classified as agranulocytes (lymphocytes and monocytes). However, both agranulocytes and granulocytes possess small number of nonspecific azurophilic granules, which are lysosomes.
, and cells that lack specific granules are classified as agranulocytes (lymphocytes and monocytes). However, both agranulocytes and granulocytes possess small number of nonspecific azurophilic granules, which are lysosomes..")
13
N EUTROPHILS Neutrophils measure 10 to 12 µm in diameter in blood smears and are obviously larger than erythrocytes. Although named for their lack of characteristic cytoplasmic staining, they are also readily identified by their multilobal nucleus; thus, they are also called polymorphonuclear neutrophils or polymorphs. Neutrophils are motile cells; they leave the circulation and migrate to their site of action in the connective tissue. Neutrophils are active phagocytes that utilize a variety of surface receptors to recognize bacteria and other infectious agents at the site of inflammation.

14
N EUTROPHIL PHAGOCYTOSIS Phagocytosed bacteria are killed within phagolysosomes by the toxic reactive oxygen intermediates produced during respiratory burst. Phagocytosed bacteria can also be killed by a diverse arsenal of oxygen-independent killing mechanisms utilizing bacteriolytic enzymes and antimicrobial peptides.

15
The cytoplasm of a neutrophil contains three kinds of granules. The different types of granules reflect the various phagocytotic functions of the cell. Specific granules (secondary granules) are the smallest granules and are at least twice as numerous as azurophilic granules. They are barely visible in the light microscope; in electron micrographs, they are ellipsoidal. Specific granules contain various enzymes as well as complement activators and other antimicrobial peptides. Azurophilic granules (primary granules) are larger and less numerous than specific granules. They arise early in granulopoiesis and occur in all granulocytes, as well as in monocytes and lymphocytes. The azurophilic granules are the lysosomes of the neutrophil and contain myeloperoxidase (MPO) (a peroxidase enzyme). Myeloperoxidase helps to generate highly reactive bactericidal hypochlorite and chloramines. Tertiary granules in neutrophils are of two types. One type contains phosphatases (enzymes that remove a phosphate group from a substrate) and is sometimes called a phosphasome. The other type contains metalloproteinases, such as gelatinases and collagenases, which are thought to facilitate the migration of the neutrophil through the connective tissue.
 are the smallest granules and are at least twice as numerous as azurophilic granules. They are barely visible in the light microscope; in electron micrographs, they are ellipsoidal. Specific granules contain various enzymes as well as complement activators and other antimicrobial peptides. Azurophilic granules (primary granules) are larger and less numerous than specific granules. They arise early in granulopoiesis and occur in all granulocytes, as well as in monocytes and lymphocytes. The azurophilic granules are the lysosomes of the neutrophil and contain myeloperoxidase (MPO) (a peroxidase enzyme). Myeloperoxidase helps to generate highly reactive bactericidal hypochlorite and chloramines. Tertiary granules in neutrophils are of two types. One type contains phosphatases (enzymes that remove a phosphate group from a substrate) and is sometimes called a phosphasome. The other type contains metalloproteinases, such as gelatinases and collagenases, which are thought to facilitate the migration of the neutrophil through the connective tissue..")
16
E OSINOPHILS Eosinophils are about the same size as neutrophils, and their nuclei are typically bilobed. Eosinophils are named for the large, eosinophilic, refractile granules in their cytoplasm. The cytoplasm of eosinophils contains two types of granules: numerous, large, elongated specific granules and azurophilic granules. Eosinophils are associated with allergic reactions, parasitic infections, and chronic inflammation. Eosinophils develop and mature in the bone marrow. Once released from the bone marrow, they circulate in peripheral blood and then migrate to the connective tissue. Eosinophils are activated by interactions with IgG, IgA, or secretory IgA antibodies.

17
B ASOPHILS Basophils are about the same size as neutrophils and are so named because the numerous large granules in their cytoplasm stain with basic dyes Often, several hundred WBCs must be examined in a blood smear before one basophil is found. Basophils are the least numerous of the WBCs, accounting for less than 0.5% of total leukocytes. The basophil cytoplasm contains two types of granules: specific granules, which are larger than the specific granules of the neutrophil, and nonspecific azurophilic granules.

18
L YMPHOCYTES Lymphocytes are the main functional cells of the lymphatic or immune system. Lymphocytes are the most common agranulocytes and account for about 30% of the total blood leukocytes. In blood smears, the mature lymphocyte approximates the size of an erythrocyte. Three functionally distinct types of lymphocytes are present in the body: T lymphocytes, B lymphocytes, and Natural killer (NK) cells.
 cells..")
19
T HE CHARACTERIZATION OF LYMPHOCYTE TYPES IS BASED ON THEIR FUNCTION, NOT ON THEIR SIZE OR MORPHOLOGY. T lymphocytes (T cells) are so named because they undergo differentiation in the thymus. Several different types of T lymphocytes have been identified: cytotoxic, helper, suppressor, and gamma/delta (γ/δ). B lymphocytes (B cells) are so named because they were first recognized as a separate population in the bursa of Fabricius in birds or bursa-equivalent organs (e.g., bone marrow) in mammals. Natural killer (NK) cells develop from the same precursor cell as B and T cells and are so named because they are programmed to kill certain types of transformed cells.
 are so named because they undergo differentiation in the thymus. Several different types of T lymphocytes have been identified: cytotoxic, helper, suppressor, and gamma/delta (γ/δ). B lymphocytes (B cells) are so named because they were first recognized as a separate population in the bursa of Fabricius in birds or bursa-equivalent organs (e.g., bone marrow) in mammals. Natural killer (NK) cells develop from the same precursor cell as B and T cells and are so named because they are programmed to kill certain types of transformed cells..")
20
M ONOCYTES Monocytes are the precursors of the cells of the mononuclear phagocytotic system. Monocytes are the largest of the WBCs in a blood smear (average diameter, 18 µm). They travel from the bone marrow to the body tissues, where they differentiate into the various phagocytes of the mononuclear phagocytotic system — that is, connective tissue macrophages, osteoclasts, alveolar macrophages, perisinusoidal macrophages in the liver (Kupffer cells), and macrophages of lymph nodes, spleen, and bone marrow among others. Monocytes remain in the blood for only about 3 days. Monocytes transform into macrophages, which function as antigen-presenting cells in the immune system. During inflammation, the monocyte leaves the blood vessel at the site of inflammation, transforms into a tissue macrophage, and phagocytoses bacteria, other cells, and tissue debris.
. They travel from the bone marrow to the body tissues, where they differentiate into the various phagocytes of the mononuclear phagocytotic system — that is, connective tissue macrophages, osteoclasts, alveolar macrophages, perisinusoidal macrophages in the liver (Kupffer cells), and macrophages of lymph nodes, spleen, and bone marrow among others. Monocytes remain in the blood for only about 3 days. Monocytes transform into macrophages, which function as antigen-presenting cells in the immune system. During inflammation, the monocyte leaves the blood vessel at the site of inflammation, transforms into a tissue macrophage, and phagocytoses bacteria, other cells, and tissue debris..")
21
THROMBOCYTES Thrombocytes are small, membrane-bounded, anucleate cytoplasmic fragments derived from megakaryocytes. Thrombocytes (platelets) are derived from large polyploid cells (cells whose nuclei contain multiple sets of chromosomes) in the bone marrow called megakaryocytes. After entry into the vascular system from the bone marrow, the platelets circulate as discoid structures about 2 to 3 m in diameter. Their life span is about 10 days. Structurally, platelets may be divided into four zones based on organization and function: The peripheral zone The structural zone The organelle zone The membrane zone
 are derived from large polyploid cells (cells whose nuclei contain multiple sets of chromosomes) in the bone marrow called megakaryocytes. After entry into the vascular system from the bone marrow, the platelets circulate as discoid structures about 2 to 3 m in diameter. Their life span is about 10 days. Structurally, platelets may be divided into four zones based on organization and function: The peripheral zone The structural zone The organelle zone The membrane zone.")
22
Platelets function in continuous surveillance of blood vessels, blood clot formation, and repair of injured tissue.

23
FORMATION OF BLOOD CELLS (HEMOPOIESIS) Hemopoiesis (hematopoiesis) includes both erythropoiesis and leukopoiesis (development of red and white blood cells, respectively), as well as thrombopoiesis (development of platelets). Blood cells have a limited life span; they are continuously produced and destroyed. The ultimate objective of hemopoiesis is to maintain a constant level of the different cell types found in the peripheral blood. Both the human erythrocyte (life span of 120 days) and the platelet (life span of 10 days) spend their entire life in the circulating blood. Leukocytes, however, migrate out of the circulation shortly after entering it from the bone marrow and spend most of their variable life spans (and perform all of their functions) in the tissues. In the adult, erythrocytes, granulocytes, monocytes, and platelets are formed in the red bone marrow; lymphocytes are also formed in the red bone marrow and in the lymphatic tissues.
 and the platelet (life span of 10 days) spend their entire life in the circulating blood. Leukocytes, however, migrate out of the circulation shortly after entering it from the bone marrow and spend most of their variable life spans (and perform all of their functions) in the tissues. In the adult, erythrocytes, granulocytes, monocytes, and platelets are formed in the red bone marrow; lymphocytes are also formed in the red bone marrow and in the lymphatic tissues..")
24
Hemopoiesis. This chart is based on the most recent concepts in hemopoiesis. It shows blood cells’ development from hemopoietic stem cells in the bone marrow to mature cells and their distribution in the blood and connective tissue compartments. In all lineages, extensive proliferation occurs during differentiation. Cytokines (including hemopoietic growth factors) may and do act individually and severally at any point in the process from the first stem cell to the mature blood or connective tissue cell.
 may and do act individually and severally at any point in the process from the first stem cell to the mature blood or connective tissue cell..")
25
Stages of erythrocytic and granular leukocytic differentiation with Romanovsky type stain. Shown here are normal human bone marrow cells as they would typically appear in a smear.

26
CONNECTIVE TISSUE PROPER Connective tissues that belong to this category are divided into two general subtypes: Loose connective tissue, sometimes called areolar tissue, and Dense connective tissue, which can be further subcategorized into two basic types based on the organization of its collagen fibers: dense irregular connective tissue and dense regular connective tissue.

27
is characterized by loosely arranged fibers and abundant cells of various types. is a cellular connective tissue with thin and relatively sparse collagen fibers. is the site of inflammatory and immune reactions. During these reactions, loose connective tissue can swell considerably. In areas of the body where foreign substances are continually present, large populations of immune cells are maintained. For example, the lamina propria, the loose connective tissue of mucous membranes, such as those of the respiratory and alimentary systems, contains large numbers of these cells. L OOSE CONNECTIVE TISSUE

28
D ENSE IRREGULAR CONNECTIVE TISSUE is characterized by abundant fibers and few cells. contains mostly collagen fibers. Cells are sparse and are typically of a single type, the fibroblast. This tissue also contains relatively little ground substance. Because of its high proportion of collagen fibers, dense irregular connective tissue provides significant strength.

29
D ENSE REGULAR CONNECTIVE TISSUE is characterized by ordered and densely packed arrays of fibers and cells. Dense regular connective tissue is the main functional component of tendons, ligaments, and aponeuroses. the fibers are arranged in parallel array and are densely packed to provide maximum strength. The cells that produce and maintain the fibers are packed and aligned between fiber bundles.

30
CONNECTIVE TISSUE FIBERS Connective tissue fibers are of three principal types. Connective tissue fibers are present in varying amounts, depending on the structural needs or function of the connective tissue. Each type of fiber is produced by fibroblasts and is composed of protein consisting of long peptide chains. The types of connective tissue fibers are: Collagen fibers Reticular fibers Elastic fibers

31
CONNECTIVE TISSUE CELLS Connective tissue cells can be resident or wandering. The cells that make up the resident cell population are relatively stable; they typically exhibit little movement and can be regarded as permanent residents of the tissue. These resident cells include: fibroblasts and a closely related cell type, the myofibroblast, macrophages, adipocytes, mast cells, and adult stem cells. The wandering cell population or transient cell population consists primarily of cells that have migrated into the tissue from the blood in response to specific stimuli. These include: lymphocytes, plasma cells, neutrophils, eosinophils, basophils, and monocytes.

Similar presentations








 –Leukocytes (white blood.>")
. Blood functions to transport oxygen, carbon dioxide,>")



>")
 and intercellular.>")




