Download presentation
Presentation is loading. Please wait.

1
Chapter 7: Membrane Structure and Function

2
The Cell Membrane It is a fluid mixture of proteins, lipids and some carbohydrates. It has 2 layers which are composed of phospholipids.

3
The Phospholipid Bilayer Both sides of the bilayer are aqueous. The phosphate part: hydrophilic and faces outward. The lipid part: hydrophobic and faces inward.

4
The Plasma Membrane Embedded within the membrane are protein molecules which selectively transport a variety of substances across the membrane.

5
The Plasma Membrane as a Fluid Mosaic The membrane is a fluid mosaic because it contains a variety of substances

6
The Evolution of the Cell Model 1925--Two Dutch Scientists E. Gorger and F. Grendel and the idea of a phospholipid bilayer. 1935--H. Davson and J. Danielli and the sandwich model. 1972--S.J. Sanger and G. Nicholson and the phospholipid bilayer with proteins dispersed throughout the bilayer.

7
1925--Dutch Scientists E. Gorger and F. Grendel--Phospholipid Bilayer Used data that analyzed components of red blood cells and determined the membranes were comprised of lipids and proteins. Concluded the membrane must be a phospholipid bilayer because it could exist as a stable boundary between two aqueous environments. The phosphate portion outward and the lipid portion inward.

8
1935--H. Davson and J. Danielli and the sandwich model Now that it was determined to be a bilayer, the question was where to place the proteins. Experimental data showed that phosphate by itself adheres less strongly to water than the surface of a biological membrane.

9
1935--H. Davson and J. Danielli and the sandwich model They proposed a “sandwich model” to account for this phenomenon. They hypothesized that the phospholipid bilayer was sandwiched between two layers of proteins.

10
1935--H. Davson and J. Danielli and the sandwich model This model held up for a long time and was seemingly supported by electron micrographs. As time went on, researchers began to notice that the different membranes found within cells didn’t fit the sandwich model.

11
1935--H. Davson and J. Danielli and the sandwich model There were different membranes with different compositions and many of the proteins had different solubilities when mixed with water. Also, there were different thicknesses between the membranes that don’t match the thickness of the plasma membrane Mitochondria 6nm vs. the 7-8nm of the plasma membrane. Also had a different appearance.

12
1935--H. Davson and J. Danielli and the sandwich model This information led to the downfall of the sandwich model. Additionally, the detail revealed by the electron microscope provided more evidence that the sandwich model was wrong. Furthermore, the proteins in the bilayer were amphipathic.

13
1972--S.J. Singer and G. Nicolson and the Fluid Mosaic Model Hypothesis that proteins were imbedded in the phospholipid bilayer. Hydrophobic regions within the bilayer. Hydrophilic regions inside and outside. This model is good because it supports what is seen with the electron microscope and is supported by freeze-fracture.

14
Freeze-Fracture and Electron Microscopy The cell preparation is frozen, fractured with a knife and then readied for electron microscopy.

15
Freeze-Fracture and Electron Microscopy This is a freeze-fracture look at the pores in the nuclear membrane. Top figure is 120,000x Bottom figure is 264,139x Images taken from Microbiology Principles and Practices, Jackie Black.

16
1972--Singer and Nicolson and the Fluid Mosaic Model The fluidity of the membrane is variable. The lipids move very rapidly. Some proteins move quickly within the membrane, some move more slowly in a seemingly coordinated fashion. Other proteins move very slowly, if at all and are anchored to the cytoskeleton.

17
The Fluid Mosaic Model of the Cell Membrane Images taken from Microbiology Principles and Practices, Jackie Black.

18
Proteins of the Bilayer Integral proteins--span completely across the membrane--called transmembrane proteins. Peripheral proteins are found on the surfaces of the membrane.

19
6 Major Functions of the Membrane Proteins. 1. Transport 2. Enzyme activity 3. Signal transduction 4. Cell to cell recognition 5. Intercellular Joining 6. Attachment to the cytoskeleton and ECM

20
Transport Protein This protein spans the membrane and provides a hydrophilic channel for particular solutes to cross. Some shuttle substances by changing shape. Others use ATP as an energy source to move substances across a membrane--Active Transport

21
Enzyme Activity Some proteins are enzymes. Sometimes they work alone. Other times they work in a team to perform and particular task.

22
Signal Transduction Some proteins (called receptors) bind with a chemical messenger (a signal) and relay information to the inside of the cell.
 bind with a chemical messenger (a signal) and relay information to the inside of the cell.")
23
Cell-Cell Recognition Some glycoproteins serve as ID tags that are recognized by other cells.

24
Intercellular Joining Membrane proteins of adjacent cells may hook together. Tight junctions and gap junctions are examples.

25
Attachment to Cytoskeleton and ECM The elements of the cytoskeleton may bond to membrane proteins helping the cell to maintain shape and location of certain membrane proteins.

26
An Example of a Transmembrane Protein Here is an example of the 2° structure of a protein and how it plays a role in the functional protein.

27
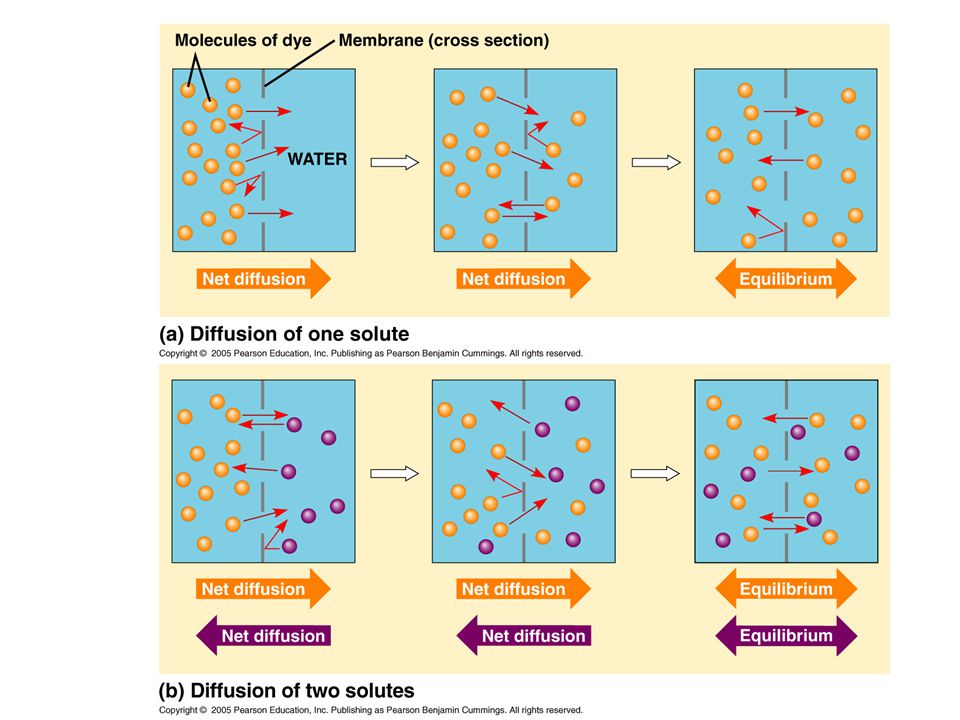
Diffusion--Passive Transport If there is a high concentration of a substance on one side of a membrane and molecules can cross the membrane, they will diffuse down their gradient until it is equal on both sides. This is passive transport.

29
Osmosis The movement of water across a selectively permeable membrane.

30
Tonicity Describes the ability of a solution to gain or lose water.

31
Tonicity Isotonic solution--no net movement of water. Hypotonic solution-- water will move into the cell. Hypertonic solution-- water will move out of the cell.

32
Tonicity It is important with animal cells because they lack a cell wall and this can cause many problems: lysis and shriveling. Not as important in plant cells because they have a cell wall which will eventually exert a back pressure slowing the uptake of water.

33
Plant Cells When the plant has more than enough water it is said to be turgid--the healthy state for non-woody plants because it provides structural support. When the plant lacks sufficient water it is flaccid. As this happens the plant begins to wilt. Plasmolysis is the dangerous state where the cell wall begins to detach from the plasma membrane.

34
Flaccid Vs. Turgid Flaccid = Wilt Turgid = Healthy

35
Facilitated Diffusion Makes no use of energy to transfer substance into and out of cells. Occurs when substances cross a membrane and diffuse down a concentration gradient.

36
2 Types of Facilitated Diffusion One type is an open channel through which substances can freely pass. The other type occurs when a protein changes shape while moving substances across a membrane.

37
Active Transport and the Na+/K+ ATPase Pump This pump operates to keep the sodium concentration low within the cell and the potassium levels high. It uses ATP as the energy to do this.

38
Exocytosis As described in Chapter 6, the trans face of the Golgi apparatus buds off a transport vesicle that moves along a microtuble to be secreted from a cell in a process called exocytosis.

39
Exocytosis In this process, as the vesicle comes in contact with the plasma membrane of the cell, the two membranes rearrange themselves and merge together. In the process, the contents of the vesicle spill out of the cell.

40
Endocytosis In the process called endocytosis, a cell pinches inward and takes up something from the outside of the cell. It works in much the same way but opposite to that of exocytosis.

41
Three Types of Endocytosis: 1. Phagocytosis 2. Pinocytosis 3. Receptor mediated endocytosis

42
Phagocytosis (Gr.-phago eating) In phagocytosis, a cell engulfs a particle by wrapping pseudopodia around it and engulfing it. The food vacuole that forms is digested by hydrolytic enzymes when it fuses with a lysosome.

43
Pinocytosis (Gr.-Pino drinking) In pinocytosis, a cell takes up fluid from the outside by pinching inward.
 In pinocytosis, a cell takes up fluid from the outside by pinching inward.")
44
Receptor Mediated Endocytosis Receptor mediated endocytosis occurs when the receptors bound to the membrane recognize and bind to ligands formed on the outside of the cell. When binding occurs, a vesicle pinches inward carrying with it molecules that the cell needs transporting them to different parts of the cell.

46
Endocytosis and Exocytosis The process of endocytosis and exocytosis are the way in which the cell continually rejuvenates the plasma membrane.

47
Receptor Mediated Endocytosis LDL has been called the "bad cholesterol". High serum LDL's go along with high serum cholesterol. However, we can reduce serum cholesterol by taking it up into cells that need it (for membranes, steroid hormone production, etc.) This requires a specific LDL receptor and a working receptor mediated endocytosis process.
 This requires a specific LDL receptor and a working receptor mediated endocytosis process..")
48
Receptor Mediated Endocytosis There are two genetic mutations that cause either no uptake of LDL receptors or uptake and accumulation of cholesterol in endosomes.

49
Receptor Mediated Endocytosis The figure at the right illustrates how LDL cholesterol is taken up by receptor mediated endocytosis. After uptake, the vesicle is metabolized and free cholesterol is available for use by the cell. www.cytochemistry.net

50
Receptor Mediated Endocytosis Cholesterol bound to LDL is taken up by cells so that it can be used in construction of membranes, etc. In this case the receptor is recycled and the ligand (LDL-cholesterol) is metabolized so the free cholesterol can be released and used by the cell. www.cytochemistry.net
 is metabolized so the free cholesterol can be released and used by the cell.")
51
Receptor Mediated Endocytosis This figure shows how LDL receptors are collecting with their ligand (LDL) in a clathrin coated pit. www.cytochemistry.net

Similar presentations














 & proteins with some carbohydrates (glycolipids.>")




 PHOSPHOLIPIDS.>")
