Download presentation
Presentation is loading. Please wait.

1
Membrane structure Membrane Transport of Small Molecules Electrical Properties of Membranes

2
Importance of cell membranes
Plasma membrane separates cellular components from the environment. Allows organelles to execute specialized functions by keeping the contents of the organelle separate from the rest of the cell. Provides boundaries that establish electrochemical gradients. These are used to make ATP and to generate nerve impulses.

3
Biological membranes are thin films composed mainly of amphipathic lipids and proteins. Most of the molecules are held together by non-covalent interactions.

4
phosphatidylcholin Polar head (phospholipid)
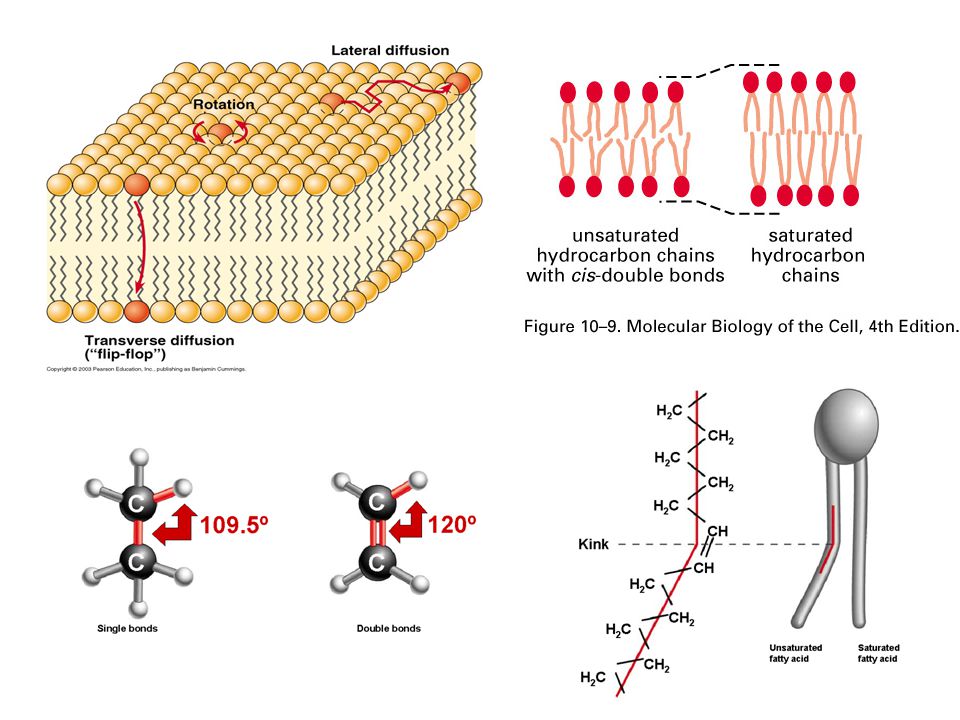
Hydrocarbon tail derived from fatty acid. Nonpolar tail Unsaturated hydrocarbon tail saturated hydrocarbon tail

5
Fatty Acids * * * * *

6
Phospholipids

7
Amphipathic molecules pack so as to minimize the interaction between water and the nonpolar parts. The two hydrocarbon tails give phospholipids a cylindrical shape that causes the molecules to pack as a bilayer in water.

9
Cholesterol Another type of lipid called cholesterol decreases fluidity because it restricts the movement of the hydrocarbon chains. Important chemical characteristics of cholesterol: Hydroxyl group constitutes the polar head group. OH is attached to rigid steroid ring. One hydrocarbon tail.

10
Cholesterol

11
Membrane fluidity

12
Fluidity of the Plasma Membrane

13
Growth of Membranes

14
FRAP (Fluorescence Recovery After Photobleaching)
")
15
FLIP (Fluorescence Loss In Photobleaching)
")
17
Membrane polarity Phosphatidylcholin Sphingomyelin Glycolipids
Cholesterol Phosphatidylserin Phosphatidylethanolamin Phosphatidylinositol

18
Membrane polarity

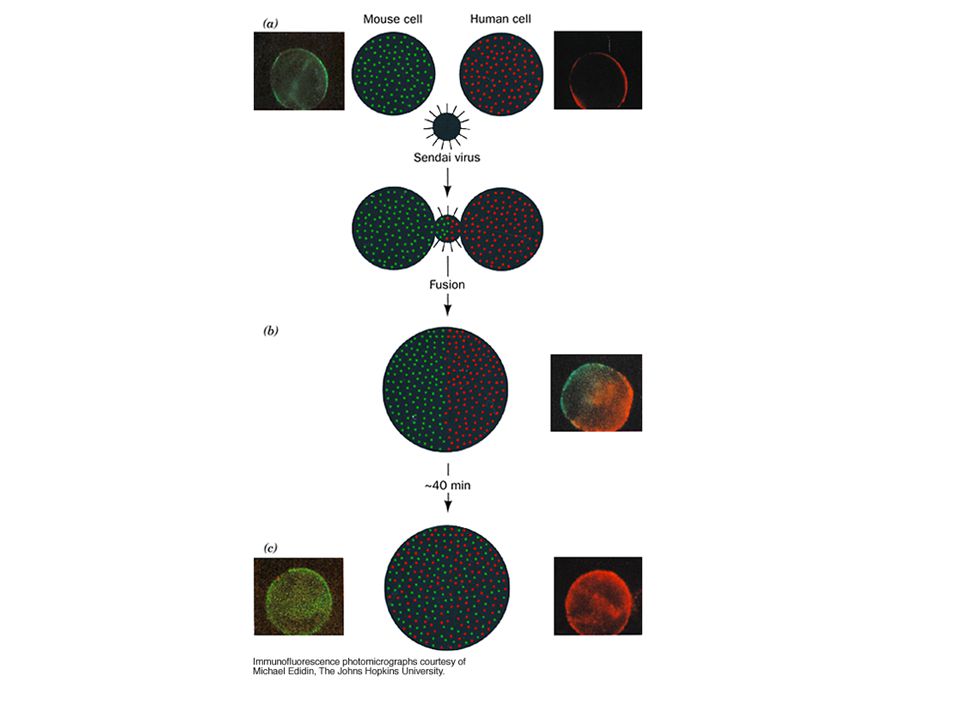
19
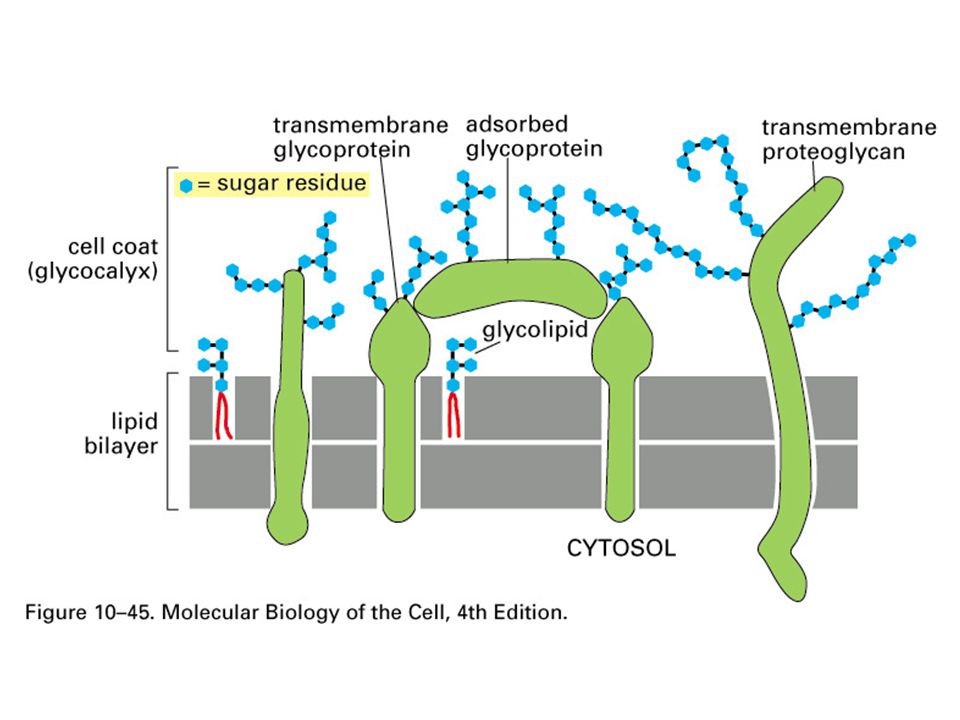
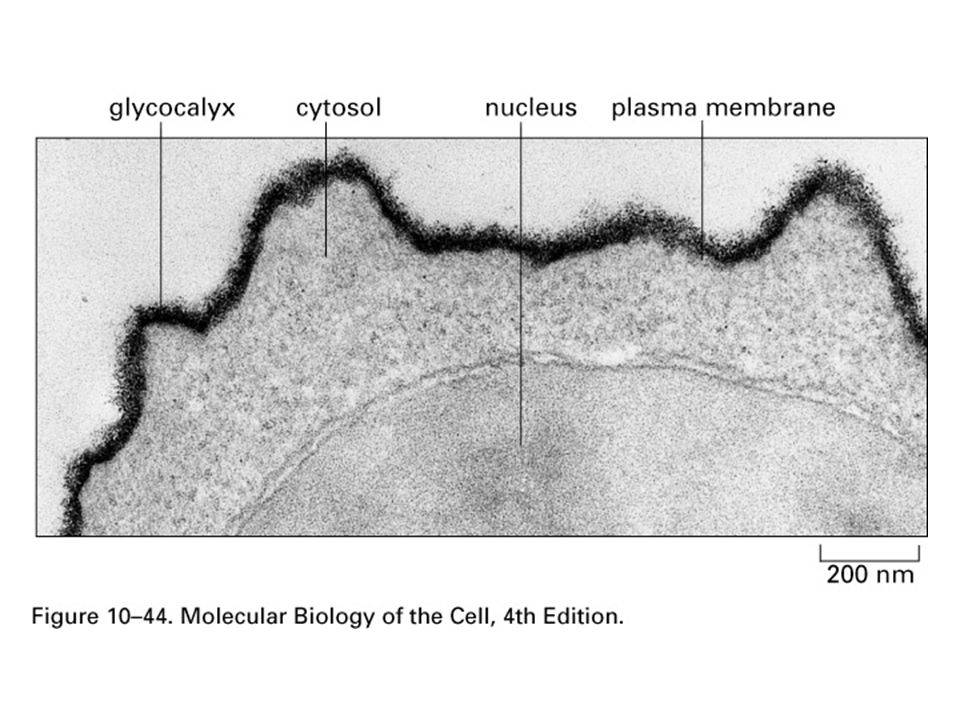
Glycolipids

22
Polarity of Plasma Membrane

23
Detergents

24
Membrane proteins Transport Cell-Cell Communication Enzymatic activity

25
Membrane proteins

26
Alpha Helix

27
Amino Acids

28
Transmembrane alpha helix
The hydrophobic core of the bilayer is 3 nm thick. One turn of the helix spans 0.54 nm and there are 3.5 a.a./turn. Hence, an transmembrane domain composed composed of an alpha-helix is around 20 amino acids long.

29
Hydropathy plots

30
Hydropathy plots

31
Glycophorin Additional characteristics of many alpha helical transmembrane proteins: SH groups facing the cytoplasm are reduced and those facing outside the cell are oxidized to S-S. Sugars are covalently attached to certain amino acid side chains in regions that face outside the cell.

32
* Protein 3 is known as a beta barrel and is composed of a beta sheet curved into a barrel Much rarer than the alpha helix transmembrane domain - limited to the outer membranes of bacteria, chloroplasts, and mitochondria. One well-known representative is a protein called Porin. The inside of the barrel is lined with polar amino acid side chains and the outside of the barrel is lined with nonpolar amino acid side chains. The polar groups in the peptide backbone are “hidden” by hydrogen bonding with antiparallel strands.

33
Beta Sheet

34
Beta Barrels transporter receptor enzyme channel

35
Detergents Mild nonionic detergent that dissolves membranes without unfolding proteins. Both are amphipathic Ionic detergent that dissolves membranes and unfolds proteins.

36
SDS-PAGE (Polyacrylamide Gel Electrophoresis)
After electrophoretic separation, proteins in the gel can be stained with a dye (Coomassie blue) which binds to protein. - Binds to most proteins (hydrophobic interactions?) in amounts roughly proportional to the molecular weight of the protein (~ 1 SDS molecule for every 2 aa residues). - Bound SDS contributes large -ve charge >> intrinsic charge of protein, so proteins are separated almost exclusively on the basis of molecular weight.
 which binds to protein. - Binds to most proteins (hydrophobic interactions ) in. amounts roughly proportional to the molecular weight of. the protein (~ 1 SDS molecule for every 2 aa residues). - Bound SDS contributes large -ve charge >> intrinsic charge. of protein, so proteins are separated almost exclusively. on the basis of molecular weight.")
37
Detergents A major advance in the study of membrane proteins involved in transport was the discovery that the proteins would spontaneously assemble with purified lipids in vesicles sometimes called “proteoliposomes”. “Liposome” indicates a vesicle made of lipids and “proteo” indicates that proteins are present.

38
Membrane proteins form complexes
eg. “photosynthetic reaction center” (from the bacterium Rhodopseudomonas) - has 4 subunits, 3 of which contain a-helical segments that span the membrane (total: 11) - helical segments are rich in non-polar aa, whose hydrophobic side chains interact with the membrane lipids that surround the complex [In red: non-protein prosthetic groups and electon- carrying isoprenoid compounds called quinones]
 - has 4 subunits, 3 of which contain. a-helical segments that span the. membrane (total: 11) - helical segments are rich in. non-polar aa, whose hydrophobic. side chains interact with the. membrane lipids that surround. the complex. [In red: non-protein prosthetic groups and electon- carrying isoprenoid compounds called quinones]")
39
Fluorescence recovery after photobleaching provides evidence that many proteins undergo lateral diffusion.

40
Cellular cortex

41
Compartmentalization of membrane proteins

42
Red Blood Cells (SEM)
")
43
The world of ghosts

44
Membrane Transport of Small Molecules

46
Permeability of Plasma Membrane

47
Transport of molecules that are impermeable to the lipid bilayer is achieved by two main classes of membrane transport proteins. Conformational change “carries” the solute across the membrane. Aqueous pore provides passage way so solute can diffuse through the membrane.

48
Four ways of molecules and ions across the membrane

49
Electrochemical gradient

50
Carrier Proteins A carrier protein binds solute on one side of a membrane, undergoes a conformational change, and releases the solute on the other side of the membrane.

51
A carrier protein resembles an enzyme
Enzyme Carrier protein Binding sites for substrates Binding sites for solute Chemically transforms substrate Transports solute Reaction rate saturate at high Transport rate saturates at high substrate concentration solute concentration

52
By coupling the conformational change to a source of energy, a carrier protein can perform active transport.

53
Depending on how many solute molecules are transported and in what direction, carrier proteins are dubbed uniporters, symporters, or antiporters.

54
Na+-K+ pump (aka Na+-K+ ATPase) in the plasma membrane is an antiporter that performs active transport. This protein establishes a concentration gradient for Na+ that is low inside the cell and high outside, and a concentration gradient for K+ this high inside the cell and low outside.

55
Direct active transport: The Na+-K+ pump (a P-class pump)
")
56
The Na+ gradient generated by the Na+ - K+ ATPase powers the transport of glucose into the cell by a Na+ -driven glucose symporter. The energetically favorable movement of Na+ down its electrochemical gradient is coupled to the energetically unfavorable transport of glucose up its concentration gradient. Hence, glucose is being subjected to active transport.

57
Glucose transport: analogy with enzyme
Sout + T k1 k-1 Sout.T k2 k-2 Sin.T k3 k-3 Sin + T SUBSTRATE PRODUCT “ENZYME” Can be described by: Kinetics: rate equations give an expression analogous to Michaelis-Menten equation: V0 = Vmax [S]out Kt + [S]out V0 = accumulation of glucose inside cell when its concentration outside is [S]out Kt = measure of affinity of transporter for S

58
Three carrier proteins, appropriately positioned in the plasma membrane, function to transport glucose across the intestinal epithelium.

59
The S.R. calcium pump

60
Osmotic pressure

61
Osmotic pressure - homeostasis
Maintaining osmotic balance is very important

62
Ion Channels and Membrane Potential

63
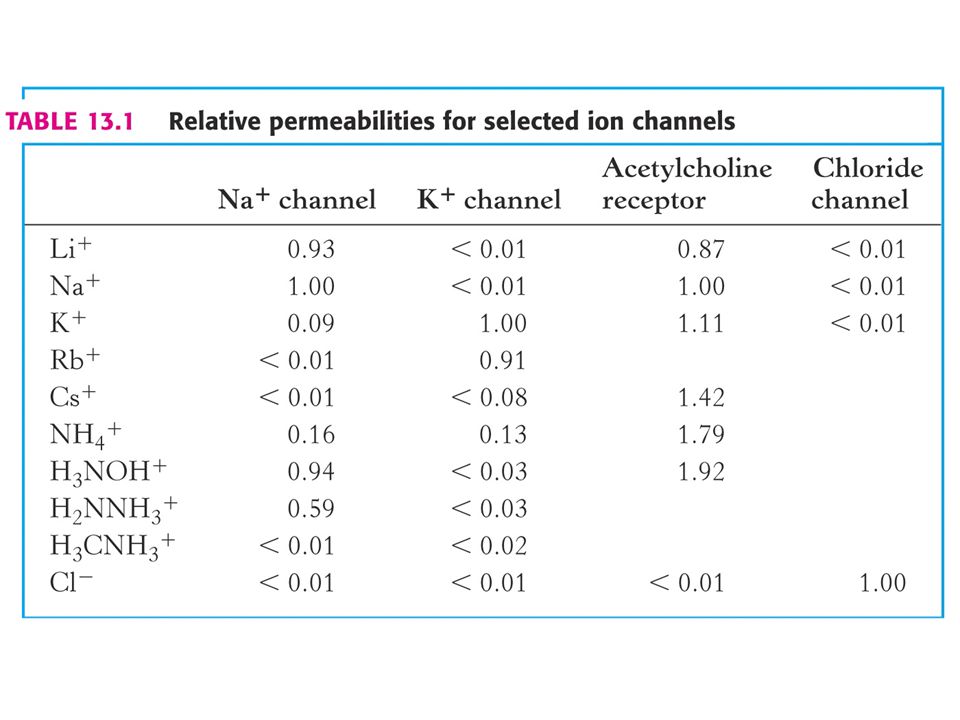
Ion channels are ion-selective because of narrow region called the “selectivity filter”.
side view R R top view - each circle represents an end-on view of an alpha-helix and the R-groups radiating outwards are amino acid side chains. R R R R R R R R R R R R R

65
Stryer Fig. 13.27 Selectivity of a sodium channel.

66
How can a K+ channel discriminate between K+ and Na+ since they both have the same charge and Na+ is smaller than K+? The answer: as the ion passes through the selectivity filter, the ion must shed water. Carbonyl oxygens with partial negative charge can take the place of water for K+ but Na+ is too small. Hence, Na+ favors remaining associated with water and hydrated ion is too large to fit through the selectivity filter.

67
Ion channels fluctuate between open and closed state to regulate ion flow.
Voltage-gated channels respond to the membrane potential. Ligand-gated channels respond to association of small molecules called ligands. Mechanically gated channels respond to movement.

68
Establishing a resting membrane potential in animal cells results from the coordinated action of carrier proteins and ion channels. 1. Na+-K+ ATPase concentrates K+ inside the cell and Na+ outside (active transport). 2. K+ leak channels allow K+ to diffuse out of the cells, down the concentration gradient (passive transport). 3. Negative charge left behind in the cytoplasm counteracts the efflux of K+ so only a very small amount (1/100,000) K+ leak out. 4. The efflux of the K+ is sufficient to generate a membrane potential of approximately -100 mV - positive outside and negative inside. The resting potential is defined as the membrane potential occurring when there is no net flow of ions. Cells typically have resting potentials between -20 and -200 mV. The major contributor is the K+ leak channel, but other channels account for the range of potentials observed.
. 2. K+ leak channels allow K+ to diffuse out of the cells, down the concentration gradient (passive transport). 3. Negative charge left behind in the cytoplasm counteracts the efflux of K+ so only a very small amount (1/100,000) K+ leak out. 4. The efflux of the K+ is sufficient to generate a membrane potential of approximately -100 mV - positive outside and negative inside. The resting potential is defined as the membrane potential occurring when there is no net flow of ions. Cells typically have resting potentials between -20 and -200 mV. The major contributor is the K+ leak channel, but other channels account for the range of potentials observed.")
69
Resting Membrane Potential: The Nernst equation

70
Nerve cell (Neuron) Dendrites - cell protrusions that receive signals from axons. Cell body - location of nucleus. Axon - single long protrusion that sends signal away from the cell body. A nerve impulse results from electrical disturbances in the plasma membrane that spread from one part of the cell to another. The electrical disturbance is called an action potential and it consists of a wave of membrane depolarization that moves down the axon.

71
Membrane depolarizations spread passively only short distances

72
Voltage-gated channels are the key to an action potential.
Depolarization to a specific membrane potential causes the channels to open. This specific membrane potential can be thought of as the threshold that must be reached to get the channel to open.

73
The structure and function of the voltage-gated Na+ channel
Figure 21-13

74
A threshold depolarization initiates the action potential.
When a nerve receives a signal, a modest depolarization of the membrane occurs. If this depolarization reaches the threshold, all the voltage-gated Na+ channels experiencing this threshold depolarization will open simultaneously. Na+ rushes into the cell causing a rapid and large depolarization of the membrane. This rapid and large depolarization is the action potential.

75
The channel cycle of closed, opened, inactivated along the axon results in propagation of the action potential.

76
Initiation of an action potential requires that something cause the membrane to depolarize to the threshold potential. The process begins at the synapse. presynaptic cell postsynaptic cell

77
Neurotransmitters are small molecules that transmit impulses at chemical synapses

78
Excitation vs. Inhibition Synapses

79
A single nerve cell combines the excitatory and inhibitory signals received from numerous other neurons to control the frequency with which action potentials are generated. The presynaptic terminals are derived from numerous other neurons. Ligand-gated ion channels are concentrated at the synapses on the postsynaptic cell (colored yellowish-brown). Neurotransmitters released into each synapse will generate a postsynaptic potential (PSP). The voltage-gated Na+ channels are concentrated at the axon hillock. An action potential is initiated at the axon hillock when the sum of excitatory and inhibitory PSP depolarize the axon hillock to the threshold potential.
. Neurotransmitters released into each synapse will generate a postsynaptic potential (PSP). The voltage-gated Na+ channels are concentrated at the axon hillock. An action potential is initiated at the axon hillock when the sum of excitatory and inhibitory PSP depolarize the axon hillock to the threshold potential.")
80
The greater the “combined PSP”, the more frequent action potentials are generated at the axon hillock. Note that the magnitude of the change in membrane potential during an action potential is constant because the membrane potential is limited by the Na+ concentration.

81
Myelination increases the speed and efficiency of action potential propagation.
Glial cells (diagrammed in red) surrounding the axon produce an electrical insulation rich in glycolipids. Nodes of Ranvier located at regular intervals are openings in the insulation where Na+ channels are concentrated. Influx of Na+ at one node results in depolarization at the next node due to the rapid diffusion of Na+ in the cytoplasm. This triggers an action potential that leads to rapid depolarization of the next node. The action potential jumps from node to node by a process called saltatory conduction.
 surrounding the axon produce an electrical insulation rich in glycolipids. Nodes of Ranvier located at regular intervals are openings in the insulation where Na+ channels are concentrated. Influx of Na+ at one node results in depolarization at the next node due to the rapid diffusion of Na+ in the cytoplasm. This triggers an action potential that leads to rapid. depolarization of the next node. The action potential jumps from node to node by a process called saltatory conduction.")
82
Action potentials travel rapidly from one node to the next
Figure 21-18

83
The neuromuscular junction is an example of how the action potential from the nerve triggers a response in another cell.

84
Electric synapses

85
Comparison of action potential transmission across electric and chemical synapses
Figure 21-36

86
The End

Similar presentations













 Nervous system functions Structure of a neuron Sensory, motor, inter- neurons Membrane potential Sodium.>")






