Download presentation
Presentation is loading. Please wait.

1
MITOCHONDRIAL DISEASES
PHM142 UNIT 7 MITOCHONDRIAL DISEASES PART 1 - Introduction

2
The mitochondrial energy-generating system

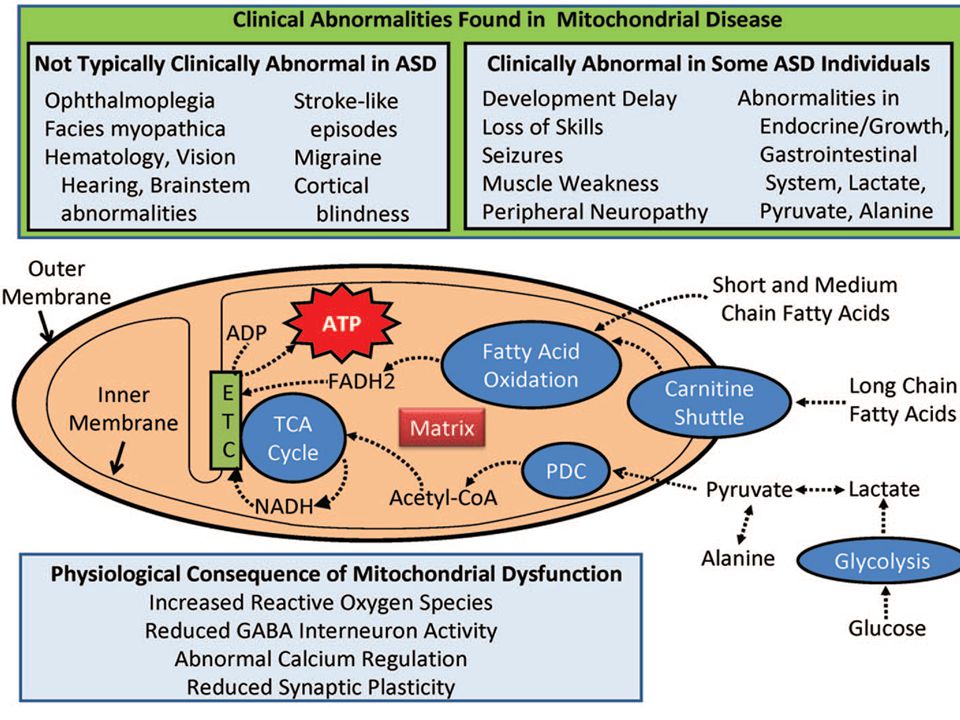
4
Mitochondrial Myopathies
Genetic defects in mitochondrial structure & function leading to defective aerobic energy transduction and resulting in: exercise intolerance, lactic acidosis, stroke/seizure, headaches.

5
Assays for mitochondrial dysfunction

7
Nijmegen Center for Mitochondrial Disorders
The table gives the percentage of fresh muscle samples that showed an activity below the lowest control value. Results from frozen muscle samples are not included in this table. In total, 1,406 fresh muscle samples were examined. Of these, 39% showed a reduced rate of ATP production from the oxidation of pyruvate and malate. Approximately 2/3 of the samples with a reduced ATP production rate also showed a reduced activity of one or more OXPHOS enzymes.

8
Mitochondrial dysfunction in neurodevelopmental disorders
PHM142 UNIT 8A Mitochondrial dysfunction in neurodevelopmental disorders (e.g. autism)
")
9
Of the 120 children with ASD studied by Oliveira, 2005 detailed metabolic studies including plasma lactate were performed in 69. Elevated lactate was found in 14, of whom 11 underwent muscle biopsy. Five of these children were diagnosed with definite mitochondrial disease. Thus, 4.2% of 120 children with ASD were determined to have definite mito disease. <> This is likely an underestimate.

11
Abnormalities in Mitochondria Function in Autism
C. Giulivi et al., JAMA 304: 2389

12
JAMA. 2010;304(21):2389-2396 Conclusions:
In this exploratory study, children with autism were more likely to have mitochondrial dysfunction, mtDNA over-replication, and mtDNA deletions than typically developing children. Using the control values to define a reference range (set at 99% CI) for the individual data, low NADH oxidase activity was the most common deficiency (8 of 10), followed by succinate oxidase (6/10). The lactate-to-pyruvate ratio reflects the redox state of the cytosolic compartment, such that a lactate-to-pyruvate ratio of 12 (as in controls) indicates a ratio of oxidized NADH to reduced NADH of 750:1, and a lactate-to-pyruvate ratio of 6 (as in autism) indicates a ratio of oxidized NADH to reduced NADH of 1500:1. A more oxidized cytosolic redox state in autism could favor anaerobic glycolysis over oxidative phosphorylation as a source of adenosine triphosphate. Although skeletal muscle can tolerate this shift in metabolism, consequences for brain function could be devastating due to its heavy reliance on mitochondrial oxidative phosphorylation to generate the energy needed for cellular processes. The rates of H2O2 production in lymphocytic mitochondria from children with autism were higher compared with controls at both complex I and complex III. Thus, lymphocytic mitochondria in autism not only had a lower oxidative phosphorylation, but also an increase in cellular oxidative stress.
 for the individual data, low NADH oxidase activity was the most common deficiency (8 of 10), followed by succinate oxidase (6/10). The lactate-to-pyruvate ratio reflects the redox state of the cytosolic compartment, such that a lactate-to-pyruvate ratio of 12 (as in controls) indicates a ratio of oxidized NADH to reduced NADH of 750:1, and a lactate-to-pyruvate ratio of 6 (as in autism) indicates a ratio of oxidized NADH to reduced NADH of 1500:1. A more oxidized cytosolic redox state in autism could favor anaerobic glycolysis over oxidative phosphorylation as a source of adenosine triphosphate. Although skeletal muscle can tolerate this shift in metabolism, consequences for brain function could be devastating due to its heavy reliance on mitochondrial oxidative phosphorylation to generate the energy needed for cellular processes. The rates of H2O2 production in lymphocytic mitochondria from children with autism were higher compared with controls at both complex I and complex III. Thus, lymphocytic mitochondria in autism not only had a lower oxidative phosphorylation, but also an increase in cellular oxidative stress.")
13
PHM 142 UNIT 8B Mitochondrial function in neurodegenerative disorders

17
- The loss of autophagy-related genes results in neurodegeneration and abnormal protein accumulation. Autophagy is a bulk lysosomal degradation pathway essential for the turnover of long-lived, misfolded or aggregated proteins, as well as damaged or excess organelles. The accumulation and aggregation of a-synuclein is a characteristic feature of PD. Over-expression of a-synuclein is thought to impair autophagy, suggesting the presence of a cycle of impairment and accumulation. Prior studies have shown that a-synuclein is degraded by chaperone-mediated autophagy. - Neurotoxins affecting humans and also used in animal models of PD: MPTP, 6-hydroxy-dopamine (6-OHDA), rotenone, and paraquat MPTP, a selective inhibitor of PD mitochondrial complex I, directed researchers’ attention to pathological roles of mitochondria in PD and raised the possibility that environmental toxins affecting mitochondria might cause PD. Other mitochondrial toxins characterized as parkinsonism-inducing reagents include 6-OHDA, rotenone, and paraquat. Studies of animal models of PD induced with these toxins suggest that mitochondrial dysfunction and oxidative stress are important pathogenic mechanisms. In humans, reduced complex I activity has been reported in both post-mortem brain samples and platelets of sporadic/idiopathic PD cases. - Lewy bodies observed in PD brain tissue are proteinaceous intracellular inclusions containing ubiquitin and a-synuclein among many other components. The protein Parkin, mutated in the most common cause of recessive PD, may mediate the clearance of abnormal mitochondria through autophagy. Recent studies have revealed that genes associated with autosomal recessive forms of PD such as PINK1 and Parkin are directly involved in regulating mitochondrial morphology and maintenance, abnormality of which is also observed in the more common, idiopathic forms of PD. Note however that the autosomal recessive PDs lack Lewy-body pathology that is characteristic of idiopathic PD.
, rotenone, and paraquat. MPTP, a selective inhibitor of PD mitochondrial complex I, directed researchers’ attention to pathological roles of mitochondria in PD and raised the possibility that environmental toxins affecting mitochondria might cause PD. Other mitochondrial toxins characterized as parkinsonism-inducing reagents include 6-OHDA, rotenone, and paraquat. Studies of animal models of PD induced with these toxins suggest that mitochondrial dysfunction and oxidative stress are important pathogenic mechanisms. In humans, reduced complex I activity has been reported in both post-mortem brain samples and platelets of sporadic/idiopathic PD cases. - Lewy bodies observed in PD brain tissue are proteinaceous intracellular inclusions containing ubiquitin and a-synuclein among many other components. The protein Parkin, mutated in the most common cause of recessive PD, may mediate the clearance of abnormal mitochondria through autophagy. Recent studies have revealed that genes associated with autosomal recessive forms of PD such as PINK1 and Parkin are directly involved in regulating mitochondrial morphology and maintenance, abnormality of which is also observed in the more common, idiopathic forms of PD. Note however that the autosomal recessive PDs lack Lewy-body pathology that is characteristic of idiopathic PD.")
18
From Fig. above Mitochondrial fusion and fission events are required for the maintenance of a healthy mitochondrial population (beige). Mitochondrial fusion is thought to facilitate the interchange of internal components such as copies of the mitochondrial genome, respiratory proteins and metabolic products. Mitochondrial fission may play a role in the removal of dysfunctional mitochondria (dark red) with reduced mitochondrial membrane potential (Dcm), through an autophagy-lysosomal pathway named ‘mitophagy’. PINK1 and Parkin are likely to be involved in this process. PINK1 normally has a short half-life in healthy mitochondria. Upon reduction of the Dcm, PINK1 is stabilized on the outer membrane; then accumulation of PINK1 induces the translocation of Parkin from the cytosol to the mitochondria, leading to Parkin-dependent ubiquitination and degradation of the mitochondrial proteins, and subsequent activation of the autophagy machinery. Ubiquitinated proteins of the mitochondria are shown as ovals with small orange circles. Recent studies of Parkin-deficient or PINK1-deficient mice have reported morpho-logical and functional alterations of mitochondria in both neurons and astrocytes . Also, a missense mutation in another gene called PARL found in PD cases abolishes its PINK1-processing activity and the ensuing Parkin-mediated mitophagy.
. Mitochondrial fusion is thought to facilitate the interchange of internal components such as copies of the mitochondrial genome, respiratory proteins and metabolic products. Mitochondrial fission may play a role in the removal of dysfunctional mitochondria (dark red) with reduced mitochondrial membrane potential (Dcm), through an autophagy-lysosomal pathway named ‘mitophagy’. PINK1 and Parkin are likely to be involved in this process. PINK1 normally has a short half-life in healthy mitochondria. Upon reduction of the Dcm, PINK1 is stabilized on the outer membrane; then accumulation of PINK1 induces the translocation of Parkin from the cytosol to the mitochondria, leading to Parkin-dependent ubiquitination and degradation of the mitochondrial proteins, and subsequent activation of the autophagy machinery. Ubiquitinated proteins of the mitochondria are shown as ovals with small orange circles. Recent studies of Parkin-deficient or PINK1-deficient mice have reported morpho-logical and functional alterations of mitochondria in both neurons and astrocytes . Also, a missense mutation in another gene called PARL found in PD cases abolishes its PINK1-processing activity and the ensuing Parkin-mediated mitophagy.")
19
Regulation of mitophagy by PINK1 and Parkin (genes linked to PD)
Current Opinion in Neurobiology 2011, 21:935–941

20
Summary a new hypothesis on mitochondrial involvement in PD
Prominent pathological features of PD include mitochondrial dysfunction and the accumulation of protein inclusions into Lewy-bodies in dopaminergic neurons. These disease phenotypes could arise from impairments in the cellular quality control systems for mitochondria and cytoplasmic proteins involving mitochondrial fission/fusion dynamics, the ubiquitin–proteasome system, and the autophagy pathway.

Similar presentations





 A 42 deposits subtle.>")



 ط Dual role of glycolysis; degrades glucose to generate ATP and source.>")

 Only found in ANIMAL cells. Mitochondria.>")












