Download presentation
Presentation is loading. Please wait.

1
MOLECULAR MECHANISMS OF HIV POSTINTEGRATION LATENCY: STRATEGIES FOR ERADICATION How is HIV transcribed? What is latency/reservoir? When is it established? How best to eradicate it Mechanistic details

2
ACTIVE REPLICATIONLATENCY

3
Nature, 1987 ELONGATION

4
SHORT AND LONG TRANSCRIPT FROM HIV LTR

5
TRANSCRIPTIONAL ELONGATION CONTROL IN PROKARYOTES AND EUKARYOTES LAMBDA N:NutB SITE vs HIV TAT:TAR RNA N-TEF: NELF and DSFI NELF: NELF-A to E (RD) DSIF = Spt4 AND Spt5 P-TEFb = CycT1 and Cdk9 NELF
 DSIF = Spt4 AND Spt5 P-TEFb = CycT1 and Cdk9 NELF")
6
NEW BIOLOGY Important role for control of elongation of transcription in eukaryotic biology: 80% untranscribed genes have paused RNA polymerase II on their promoters Introduced positive and negative elongation factors to transcription (P-TEFb, N-TEF): revealed that they play the key role in HIV replication: epigenetic changes result from the interplay between negative and positive elongation factors on RNA polymerase II on promoters Introduced the control of co-transcriptional processing of RNA, i.e. capping, splicing and polyadenylation: extent of RNA polymerase II phosphorylation dictates alternative splicing, which includes the processing of HIV genomic RNA Provided the first glimpse of HIV proviral latency in the host: stalled RNA polymerase II on inactive proviruses

7
PCR AMPLIFICATION OF SHORT AND LONG TRANSCRIPTS

8
Adams et al., PNAS, 1994, 10 individuals, not on HAART, at seroconversion had only short transcripts in the periphery-proviral latency (replication elsewhere)
")
9
HIV Latency HIV can hide in very long lived memory CD4 T cells- insensitive to antiviral drugs which require viral replication While these cells are rare (10 5 -10 6 /pt), clearing them from the body is predicted to require >60 years of treatment New approaches are urgently needed to purge or inactivate these latent proviruses, otherwise a “cure” for HIV infection will remain out of reach
, clearing them from the body is predicted to require >60 years of treatment New approaches are urgently needed to purge or inactivate these latent proviruses, otherwise a cure for HIV infection will remain out of reach")
10
HIV LTR RNAPII S5 S2 Valproic acid HIV LTR RNAPII S5 S2 HDAC1 RNAPII S5 S2 A B P50 HDAC1 Nuc-(+1) CHROMATIN EFFECTS
 CHROMATIN EFFECTS")
11
HIV LTR P50 RelA IBIB Prostratin, TNF HIV LTR RNAPII S5 S2 RNAPII S5 S2 P50 RelA A B Nuc-(+1) RNAPII S5 S2 RNAPII S5 S2 HATs LACK OF NF- B
 RNAPII S5 S2 RNAPII S5 S2 HATs LACK OF NF- B")
12
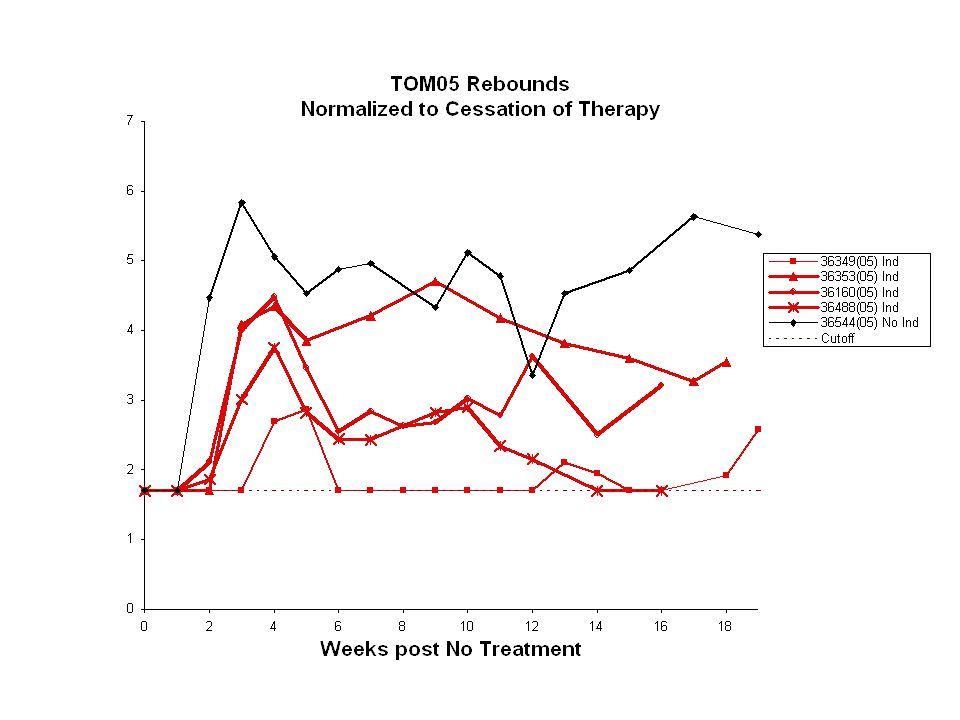
Experimental plan: RT-SHIV and HAART and PRS + VPA Induction 6 wks32 - 35 wks 8 wks 1 wk RT-SHIV Inoculation (6) HAART (FTC + PMPA + Efavirenz) HAART + Induction Necropsy (2) Rebound (4) HAART 2 wks: 5 cycles 6 wks: 1 cycle per wk Weekly/biweekly analysis: plasma viral RNA, CBC, FACS
 HAART (FTC + PMPA + Efavirenz) HAART + Induction Necropsy (2) Rebound (4) HAART 2 wks: 5 cycles 6 wks: 1 cycle per wk Weekly/biweekly analysis: plasma viral RNA, CBC, FACS")
14
HIV LTR RNAPII S5 S2 RNAPII S5 S2 P-TEFb Tat A B RNAPII S5 S2 RNAPII S5 S2 LACK OF TAT/P-TEFb

15
HIV 5’ LTRHIV 3’ LTRHost gene promoter RNAPII S5 S2 RNAPII S5 S2 polyA signal pA RNAPII S5 S2 RNAPII S5 S2 RNAPII S5 S2 HIV 5’ LTRHIV 3’ LTRHost gene promoter RNAPII S5 S2 polyA signal pA RNAPII S5 S2 RNAPII S5 S2 Activation of HIV 5’LTR (NF- B) with Prostratin, TNF A B RNAPII S5 S2 TRANSCRIPTIONAL INTERFERENCE
 with Prostratin, TNF A B RNAPII S5 S2 TRANSCRIPTIONAL INTERFERENCE")
16
Tat Cdk9 CycT1 Cdk9 RelA p50 CycT1 Cdk9 CycT1 Cdk9 activation binding direct exchange HEXIM1 7SK RNA TAR RNA BRD4 NF- B TRANSCRIPTION FROM HIV LTR DEPENDS ON P-TEFb STRESS, UV LIGHT DRB, ACTINOMYCIN D, CELL SIGNALING HMBA, SAHA, ETC.

17
PARADOX: HMBA INDUCES HEXIM1/2 BUT IS ALSO ONE OF THE MOST POTENT ACTIVATORS OF HIV TRANSCRIPTION, EVEN FROM LATENTLY INFECTED CELL LINES. HMBA IS ALSO ONE OF THE MOST POTENT CELLULAR DIFFERENTIATION AGENTS Cdk9 CycT Active P-TEFb HEXIM1/2 7SK snRNA Inactive P-TEFb LARP7 MePCE HOW TO ACTIVATE HIV TRANSCRIPTION FROM LATENCY?

18
HMBA releases P-TEFb from the LC HMBA LC SC 1h2h- HMBA 30’6h24h Active P-TEFb

19
0 3 6 12 24 hrs 100% 50% inactive P-TEFb levels of HEXIM1/2 Effects of HMBA on P-TEFb and HEXIM1/2

20
HMBA PI3K PIP2 PDK1/2 P cytoplasm nucleus HIV LTR CycT1 7SK snRNA P Inactive P-TEFb Active P-TEFb PIP3 AKT HEXIM1 P CDK9 transcriptional elongation 7SK snRNA P RNAPIIo 5 2 P

22
SAHA activates the Akt pathway Liu et al. JBC 2006 P-TEFb

23
HMBA/SAHA reactivate HIV in PBMCs Infection in vitroPBMCs from HAART treated patients PBMCs from healthy donors+PHA/IL2 Infection in vitro HIV-LAI Treatment with SAHA/HMBA 3 days 11 days PBMCs + SAHA/HMBA 2/3 days Wash+Add PBMCs treated PHA/IL2 From healthy donors P24 ELISA 15 to 21 days Negative Control SAHAAI8 + SAHA PHA + IL-2 -+-+

25
SAHA

26
SAHA (+/- NF- B ACTIVATION) Rhesus macaque trial with SAHA (+/- Prostratin) (NF- B activation will counteract chromatin effects and transcriptional interference and recruit more RNA polymerase II to the HIV LTR) SAHA will release and activate P-TEFb transiently from the large, inactive complex This manipulation will lead to the synthesis of Tat, which can utilize P-TEFb from the inactive complex to sustain HIV replication Increased levels of HEXIM1/2 and inactive complex will inhibit other viruses (HSV, EBV, CMV, KSVH), attenuate immune activation and help prevent superinfection by HIV.
 Rhesus macaque trial with SAHA (+/- Prostratin) (NF- B activation will counteract chromatin effects and transcriptional interference and recruit more RNA polymerase II to the HIV LTR) SAHA will release and activate P-TEFb transiently from the large, inactive complex This manipulation will lead to the synthesis of Tat, which can utilize P-TEFb from the inactive complex to sustain HIV replication Increased levels of HEXIM1/2 and inactive complex will inhibit other viruses (HSV, EBV, CMV, KSVH), attenuate immune activation and help prevent superinfection by HIV.")
27
Matjaz Barboric, Koen Bartholomeeusen, Dalibor Blazek, Xavier Contreras, Jiri Kohoutek, Audrey Low, Tina Lenasi, Fan Zhang COLLABORATION: PPG: Verdin, Greene, Peterlin Steve Deeks, Su Guo, Paul Luciw, Jeffrey Martin, Thomas North

Similar presentations










 Nat Rev Immunol. 12(8): 607-614.>")










