Download presentation
Presentation is loading. Please wait.

1
MEDICAL REHABILITATION EMBRYOLOGICAL DEVELOPMENT OF THE SKELETAL SYSTEM

2
INTRODUCTION TO : The Embryology of the Skeletal System The skeletal system develops from paraxial and lateral plate (somatic layer) mesoderm and from neural crest. Paraxial mesoderm forms a segmented series of tissue blocks on each side of the neural tube, known as somitomeres in the head region and somites from the occipital region caudally.

3
Somites differentiate into a ventromedial part, the sclerotome, and a dorsolateral part, the dermomyotome. At the end of the fourth week sclerotome cells become polymorphous and form a loosely woven tissue, the mesenchyme, or embryonic connective tissue. It is characteristic for mesenchymal cells to migrate and to differentiate in many ways. They may become fibroblasts, chondroblasts, or osteoblasts (bone-forming cells).
..")
4
The bone-forming capacity of mesenchyme is not restricted to cells of the sclerotome, but occurs also in the somatic mesoderm layer of the body wall, which contributes mesoderm cells for formation of the pelvic and shoulder girdles and the long bones of the limbs. Neural crest cells in the head region also differentiate into mesenchyme and participate in formation of bones of the face and skull.

5
Occipital somites and somitomeres also contribute to formation of the cranial vault and base of the skull. In some bones, such as the flat bones of the skull, mesenchyme in the dermis differentiates directly into bone, a process known as intramembranous ossification. In most bones, however, mesenchymal cells first give rise to hyaline cartilage models, which in turn become ossified by endochondral ossification.

6
SKULL The skull can be divided into two parts: the neurocranium, which forms a protective case around the brain, and the viscerocranium, which forms the skeleton of the face. The neurocranium is most conveniently divided into two portions: (a) the membranous part, consisting of flat bones, which surround the brain as a vault; and(b) the cartilaginous part, or chondrocranium, which forms bones of the base of the skull.
 the membranous part, consisting of flat bones, which surround the brain as a vault; and(b) the cartilaginous part, or chondrocranium, which forms bones of the base of the skull..")
7
NEWBORN SKULL At birth the flat bones of the skull are separated from each other by narrow seams of connective tissue, the sutures, which are also derived from two sources: neural crest cells (sagittal suture) and paraxial mesoderm (coronal suture). At points where more than two bones meet, sutures are wide and are called fontanelles. The most prominent of these is the anterior fontanelle, which is found where the two parietal and two frontal bones meet. Sutures and fontanelles allow the bones of the skull to overlap (molding) during birth.
 during birth..")
8
CLINICAL IMPLICATIONS Cranial defects and skeletal dysplasias Neural crest cells- these originate in the neuroectoerm from facial skeleton and most of the skull. Craniosis- the cranial vault fails to form cranioschsis meaning that the brain tissue exposed to amniotic fluid degenerates ( anencephaly). Microcephaly- the brain fails to grow and the skull fails to expand. Craniosynostosis and dwarfism
. Microcephaly- the brain fails to grow and the skull fails to expand. Craniosynostosis and dwarfism.")
9
THIS IS CAUSED BY HYPERPITUTARISM AND PRODUCTION OF GROWTH HORMONE ACROMEGALY

11
LIMB GROWTH AND THE DEVELOPMENT OF THE FETUS After four weeks of development, the buds for the limbs become visible from the ventrolateral body wall. It consists of a mesenchymal core that will form the bones and connective tissues of the limb which is covered by a layer of cuboidal ectoderm. The ectoderm at the distal border of the limb then becomes thicker which forms after the apical ectodermal ridge (AER). As the limbs continue growing, the cells that are farther away from, the influence of AER begins to form into muscles and cartilage.
. As the limbs continue growing, the cells that are farther away from, the influence of AER begins to form into muscles and cartilage..")
12
5 WEEKS 6 WEEKS 8 WEEKS THE LIMB BUDS DEVELOP AS THE EMBRYO GROWS

13
The limb buds of the six week embryo now become flattened to form the hand plates and foot plates which is separated by a circular constriction. Fingers and toes are formed.

14
Development of the upper and lower limb is similar except that the morphogenesis of the lower limb is approximately one to two days behind that of the upper limbs. During the seventh week of gestation, the limbs (both upper and lower) rotate in opposite directions. The upper limbs rotates ninety degrees laterally so that the extensor muscles lie on the lateral posterior surface and the thumbs lie laterally, meanwhile the lower limbs rotates ninety degrees medially placing the extensor muscles on an anterior surface and the big toe medially.
 rotate in opposite directions. The upper limbs rotates ninety degrees laterally so that the extensor muscles lie on the lateral posterior surface and the thumbs lie laterally, meanwhile the lower limbs rotates ninety degrees medially placing the extensor muscles on an anterior surface and the big toe medially..")
15
By the sixth week of development of the embryo the first hyaline cartilage models are formed by chondrocytes. Joints are now formed in the cartilaginous condensations. Cells in this region then increase in number and in density and join a cavity that is formed by cell death. Ossification of the bones of the extremities, endochondral ossification begins by the end of the embryonic period. The primary ossification centers are present in all of the long bones of the limbs of the embryo by the twelfth week of development.

16
At birth the diaphysis of the bones are usually completely ossified but the two ends; the epiphyses are still cartilaginous. A temporary a cartilage plate remains between the diaphyseal epiphyseal ossification centers. The epiphyseals plate plays an important role in growth and length of the bones of the embryo. When the bones has acquired its full length, the epiphyseal plates disappear and the epiphyses then unite with the shaft of the bone. In long bones an epiphyseal plate is formed in each extremity in smaller bones, such as the phalanges; it is found only at one extremity and in irregular bones such as the vertebrae one or more primary centers of ossification.

17
Positioning of the limbs along the cranio - caudal axis in the flank regions of the embryo is regulated by the HOX genes expressed along this axis. Thesehomeoboxgenes are expressed in overlapping patterns from head to tail, with some having more cranial limits than others. For example, thecranial limit of expression of HOXB8 is at the cranial border of the forelimb,andmisexpression of this gene alters the position of these limbs

18
Although patterning genes for the limb axes have been determined, it is the HOX genes that regulate the types and shapes of the bones of the limb. Thus, HOX gene expression occurs in phases in three places in the limb that correspond to formation of the proximal (stylopod), middle (zeugopod), and distal (autopod) parts. Genes of the HOXA and HOXD clusters are the primary determinants in the limb, and variations in their combinatorial patterns of expression may account for differences in forelimb and hindlimbstructures. Just as in the craniocaudal axis of the embryo, HOX genes are nested in overlapping patterns of expression that somehow regulate patterning.
, middle (zeugopod), and distal (autopod) parts. Genes of the HOXA and HOXD clusters are the primary determinants in the limb, and variations in their combinatorial patterns of expression may account for differences in forelimb and hindlimbstructures. Just as in the craniocaudal axis of the embryo, HOX genes are nested in overlapping patterns of expression that somehow regulate patterning..")
19
CLINICALN IMPLICATION: Congenital hip dislocation consists of underdevelopment of the acetabulum and head of the femur. It is rather common and occurs mostly in females. Although dislocation usually occurs after birth, the abnormality of the bones develops prenatally. Since many babies with congenital hip dislocation are breech deliveries, it has been thought that breech posture may interfere with development of the hip joint. It is frequently associated with laxity of the jointcapsule.

20
Vertebral Column It is during the fourth week of development that cells of the sclerotomes shift their position to surround both the spinal cord and the notochord. This mesenchymal column retains traces of its segmental origin, as the sclerotomicblocks are separated by less dense areas containing intersegmental arteries. During further development the caudal portion of each sclerotome segment proliferates extensively and condenses. This proliferation is so extensive that it proceeds into the subjacent intersegmental tissue and binds the caudal half of one sclerotome to the cephalic half of the subjacent sclerotome. Hence, by incorporation of the intersegmentaltissue into the precartilaginous vertebral body, the body of the vertebra becomes intersegmental. Patterning of the shapes of the different vertebra is regulated by HOX genes.

21
Development of spinal cord in embryonic period

22
Mesenchymal cells between cephalic and caudal parts of the original sclerotome segment do not proliferate but fill the space between two precartilaginous vertebral bodies. In this way they contribute to formation of the intervertebral disc. Although the notochord regresses entirely in the region of the vertebral bodies, it persists and enlarges in the region of the intervertebral disc. Here it contributes to the nucleus pulposus, which is later surrounded by circular fibers of the annulus fibrosus. Combined, these two structures form The intervertebral disc

23
Rearrangement of sclerotomes into definitive vertebrae causes the myotomes to bridge the intervertebral discs, and this alteration gives them the capacity to move the spine. For the same reason, intersegmental arteries, at first lying between the sclerotomes, now pass midway over the vertebral bodies. Spinal nerves, however, come to lie near the intervertebral discs and leave the vertebral column through the intervertebral foramina

24
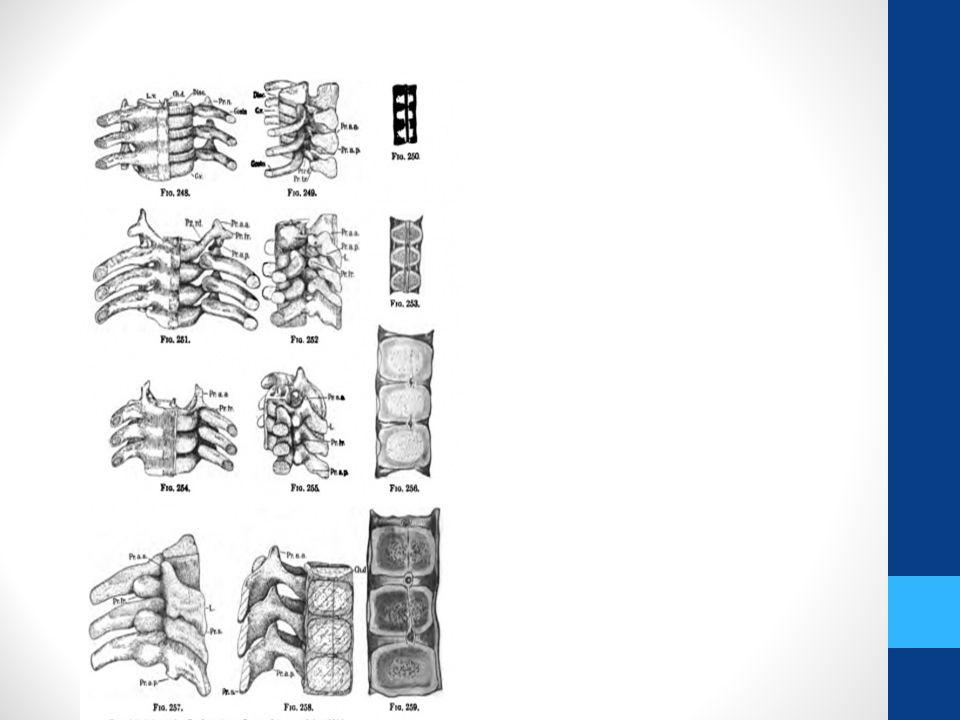
Vertebral Development

26
Development of Ribs and Sternum Ribs form from the costal processes of thoracic vertebrae and are derived from The sclerotome portion of paraxial mesoderm. The sternum develops separately in somatic mesoderm in the ventral body wall. Two sternal bands are formed on either side of the midline, and these later fuse to form cartilaginous models of the manubrium, sternebrae, and xiphoid process

27
Illustration showing the manner in which each vertebral centrum is developed from portions of two adjacent segments. (Bartleby)
.")
Similar presentations