Download presentation
Presentation is loading. Please wait.

1
The Adaptive Immune Response

2
FUNCTIONS OF THE IMMUNE SYSTEM: Recognize, destroy and clear a diversity of pathogens. Initiate tissue and wound healing processes. Recognize and clear damaged self components. Exhibit “tolerance” to innocuous material including self

3
The Normal Immune Response The normal immune response is best understood in the context of defense against infectious pathogens, the classical definition of immunity. Innate immunity refers to defense mechanisms that have evolved to specifically recognize microbes and protect individuals against infections. Adaptive immunity consists of mechanisms that are stimulated and are capable of recognizing microbial and nonmicrobial substances.

4
Innate immunity The first line of defense always ready to prevent and eradicate infections. Adaptive immunity Develops later, after exposure to microbes, and is even more powerful than innate immunity in combating infections. By convention, the term “immune response” refers to adaptive immunity.

5
Innate and Adaptive immunity represent two different arms of the immune system that work together in host defense. Innate Immunity (natural/native): Provides immediate protection from infection. Is broadly specific to microbes and tissue damage products. Does not change in response to reinfection (nonadaptive) Initiates processes that lead to activation of adaptive immune responses. Adaptive Immunity (specific/acquired): Appears to adapt to a variety of non-self components (acquired) Is highly specific to a particular molecule “antigen” Responses upon reinfection are faster, better and stronger (memory) Generates proteins and cells that enhance innate immune function.
: Provides immediate protection from infection. Is broadly specific to microbes and tissue damage products. Does not change in response to reinfection (nonadaptive) Initiates processes that lead to activation of adaptive immune responses. Adaptive Immunity (specific/acquired): Appears to adapt to a variety of non-self components (acquired) Is highly specific to a particular molecule antigen Responses upon reinfection are faster, better and stronger (memory) Generates proteins and cells that enhance innate immune function..")
6
The innate immune system provides immediate protection. The adaptive response takes time to develop and is antigen specific. Activation of B and T lymphocytes Plasma cellsNaive

7
ADAPTIVE IMMUNITY The adaptive immune system consists of lymphocytes and their products, including antibodies. The receptors of lymphocytes are much more diverse than those of the innate immune system, but lymphocytes are not inherently specific for microbes, and they are capable of recognizing a vast array of foreign substances. http://www.pathologystudent.com/wp- content/uploads/2010/07/normal-lymphs.jpg

8
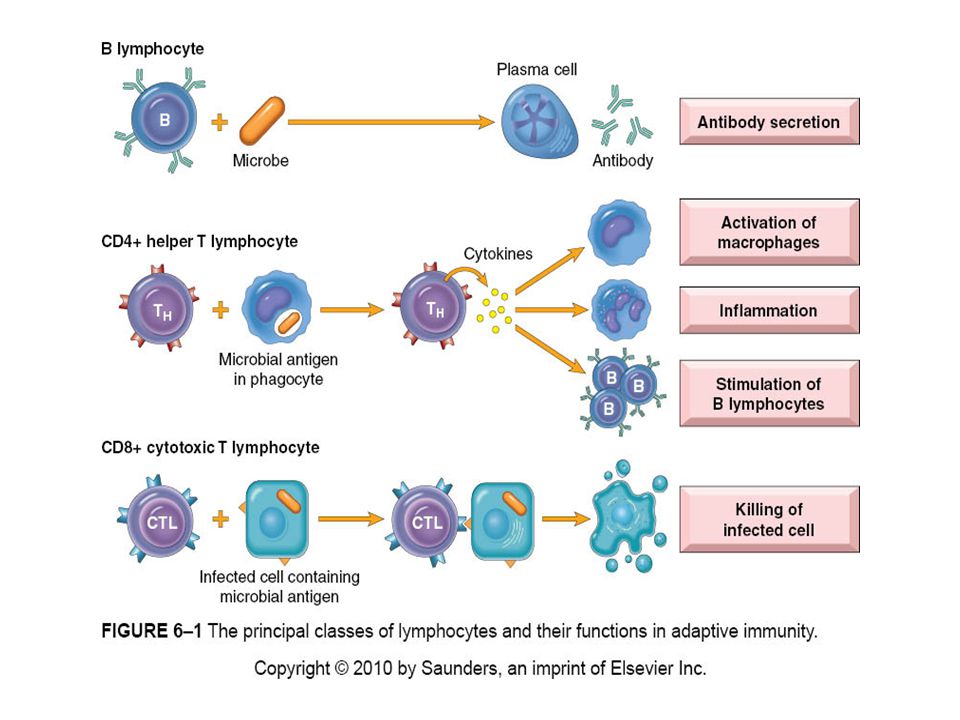
There are two types of adaptive Immunity Humoral immunity: mediated by B lymphocytes and their secreted products, antibodies (also called immunoglobulins, Ig) that protects against extracellular microbes and their toxins. Cellular immunity : mediated by T lymphocytes and is responsible for defense against intracellular microbes. http://yang-sheng.com/wp-content/uploads/2011/05/adoptive_immunity.gif

9
Types of Adaptive Immune Reponses

10
Although lymphocytes appear morphologically unimpressive and similar to one another, they are actually remarkably heterogeneous and specialized in molecular properties and functions. Lymphocytes and other cells involved in immune responses are not fixed in particular tissues (as are cells in most of the organs of the body) but are capable of migrating among lymphoid and other tissues and the vascular and lymphatic circulations. This feature permits lymphocytes to home to any site of infection. In lymphoid organs, different classes of lymphocytes are anatomically segregated in such a way that they interact with one another only when stimulated to do so by encounter with antigens and other stimuli. Lymphocytes http://www.nature.com/nri/journal/v12/n1/images/nri3128-f5.jpg
 but are capable of migrating among lymphoid and other tissues and the vascular and lymphatic circulations. This feature permits lymphocytes to home to any site of infection. In lymphoid organs, different classes of lymphocytes are anatomically segregated in such a way that they interact with one another only when stimulated to do so by encounter with antigens and other stimuli. Lymphocytes")
12
Mature lymphocytes that have not encountered the antigen for which they are specific are said to be naive (immunologically inexperienced). After they are activated by recognition of antigens and other signals lymphocytes differentiate into: effector cells, which perform the function of eliminating microbes. memory cells, which live in a state of heightened awareness and are better able to combat the microbe in case it returns. http://www.nature.com/ni/journal/v5/n9/images/ni0904-873-F1.jpg

13
http://www.nature.com/nri/journal/v7/n12/images/nri2204-f1.jpg

14
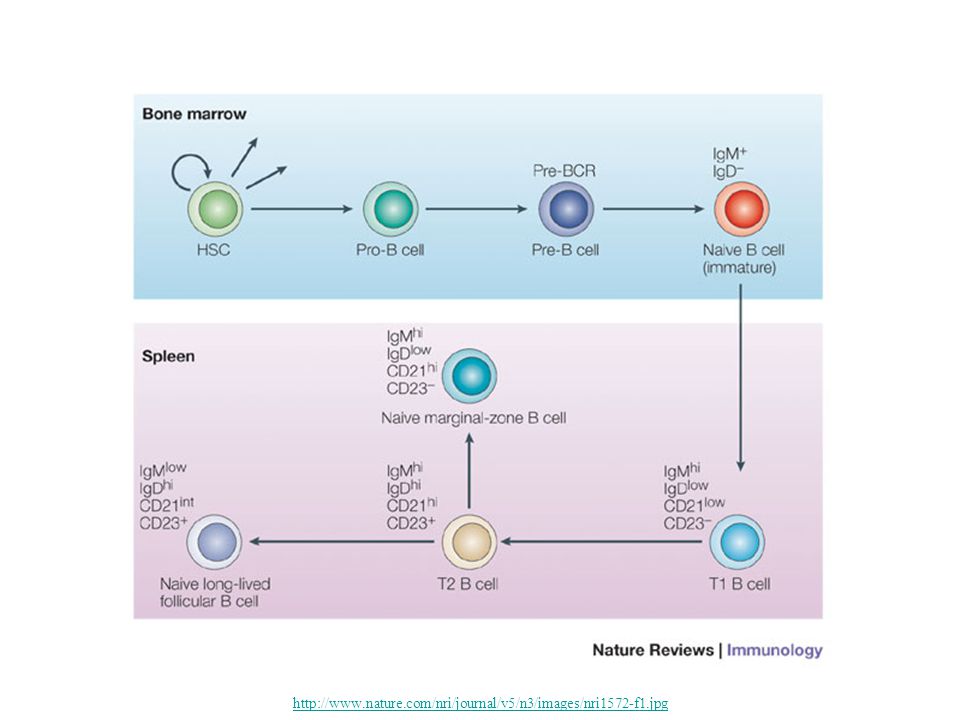
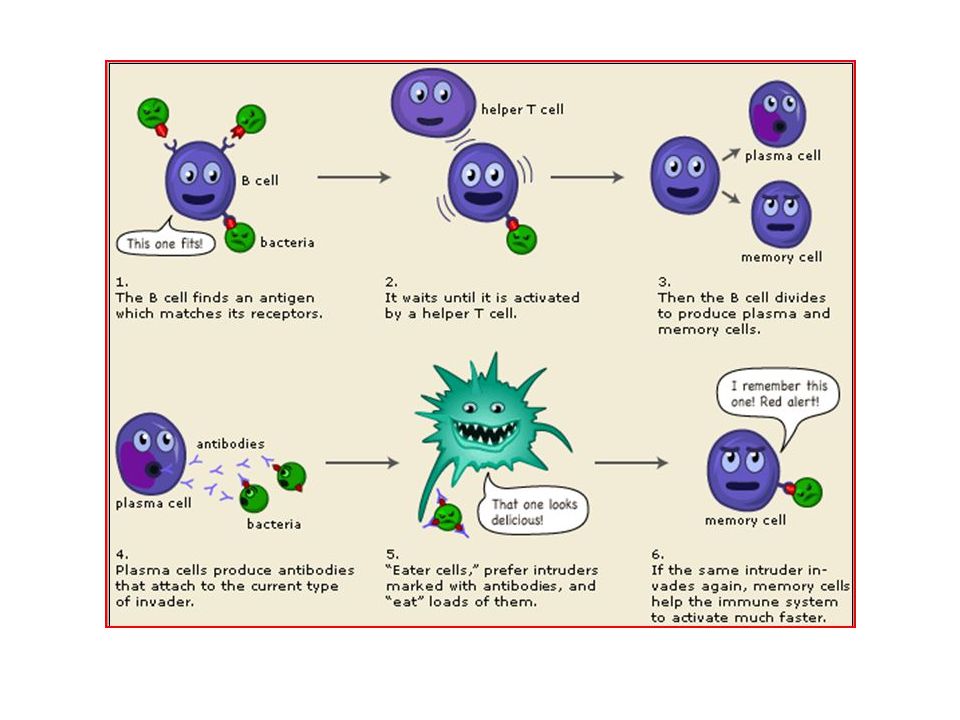
B Lymphocytes B lymphocytes develop from precursors in the bone marrow. Mature B cells constitute 10% to 20% of the circulating peripheral lymphocyte population and are also present in peripheral lymphoid tissues such as lymph nodes, spleen, and mucosa-associated lymphoid tissues. B cells recognize antigen via the B-cell antigen receptor complex. Membrane-bound antibodies called IgM and IgD, present on the surface of all mature, naive B cells, are the antigen-binding component of the B-cell receptor complex. Each B-cell receptor has a unique antigen specificity, derived from RAG- mediated rearrangements of Ig genes. After stimulation by antigen and other signals, B cells develop into plasma cells that secrete antibodies, the mediators of humoral immunity. http://theadventuresofbecky.files.wordpress.com/ 2010/03/plasma-cell-100x-website-arrow.jpg http://cytochemistry.net/cell- biology/medical/06_016.jpg

15
http://www.nature.com/nri/journal/v5/n3/images/nri1572-f1.jpg
17
Immunoglobulin S-S COOH ClVl VhCh1Ch2Ch3 S-S

18
Immunoglobulins (cont.) Biologic properties of immunoglobulin domains Ch3 - cytotrophic reactions involving: macrophages and monocytes heterologous mast cells cytotoxic killer cells B cells Ch2 binding complement control of catabolic rate Vh/Vl antigen binding
 Biologic properties of immunoglobulin domains Ch3 - cytotrophic reactions involving: macrophages and monocytes heterologous mast cells cytotoxic killer cells B cells Ch2 binding complement control of catabolic rate Vh/Vl antigen binding")
19
B Lymphocytes.. 3' (DNA) 2 4 1 3 1 JJJVVV DDD Germline Rearranged VVVD DJJ VVV VDJ J DJJ 1° Transcript VDJ µ - mRNA 2
 2 4 1 3 1 JJJVVV DDD Germline Rearranged VVVD DJJ VVV VDJ J DJJ 1° Transcript VDJ µ - mRNA 2.")
20
B Lymphocytes. Germline DNA J Hind III µ3' µ VDJ B cell leukemia µ DJ Allele A Allele B GermlineLeukemia B A Normal J probe

21
http://4.bp.blogspot.com/- IXXEpj_bFg8/T_QO506s1lI/AAAAAAAAAEc/pEUu2wprOhU/s1600/ig+ge ne+in+b+cell+differentiation.png

22
In addition to membrane Ig, the B-cell antigen receptor complex contains a heterodimer of two invariant proteins called Igα and Igβ. B cells also express several other molecules that are essential for their responses. These include complement receptors, Fc receptors, and CD40. http://www.cellsignal.com/reference/pathway/images/B_Cell_Receptor.jpg http://www.nature.com/nri/journal/v2/n2/images/nri726-f1.gif

24
Upon activation, B lymphocytes proliferate and then differentiate into plasma cells that secrete different classes of antibodies with distinct functions (Fig. 6-12).
..")
25
Many polysaccharide and lipid antigens have multiple identical antigenic determinants (epitopes) that are able to engage many antigen receptor molecules on each B cell and initiate the process of B-cell activation. Typical globular protein antigens are not able to bind to many antigen receptors, and the full response of B cells to protein antigens requires help from CD4+ T cells.

26
http://upload.wikimedia.org/wikipedia/commons/thumb/f/f7/B_cell_activation.png/300px-B_cell_activation.png B cells ingest protein antigens into vesicles, degrade them, and display peptides bound to MHC molecules for recognition by helper T cells. The helper T cells express CD40L and secrete cytokines, which work together to activate the B cells.

27
Immunoglobulins IgG, IgM and IgA constitute 95% of serum immunoglobulins IgE trace, IgD B cell membrane associated Monomeric IgM on B cell surface constitutes B cell receptor B cell antigen receptor complex contains nonpolymorphic transmembrane proteins like the CD3 proteins of TCR, Ig and Ig

28
Each plasma cell secretes antibodies that have the same antigen binding site as the cell surface antibodies (B-cell receptors) that first recognized the antigen. Polysaccharides and lipids stimulate secretion mainly of IgM antibody. Protein antigens, by virtue of CD40L- and cytokine-mediated helper T-cell actions, induce the production of antibodies of different classes, or isotypes (IgG, IgA, IgE). Cytokines that induce isotype switching include IFN-γ and IL-4.
. Cytokines that induce isotype switching include IFN-γ and IL-4..")
29
Helper T cells also stimulate the production of antibodies with high affinities for the antigen. This process, called affinity maturation, improves the quality of the humoral immune response. Isotype switching and affinity maturation occur mainly in germinal centers, which are formed by proliferating B cells, especially in helper T cell- dependent responses to protein antigens. http://www.cs.unm.edu/~immsec/html-imm/AFF-MAT1.JPG

30
Signal 1 Signal 2 Helper T cells are required for efficient isotype switching, affinity maturation and generation of long lived memory B cells

31
Antibodies bind to microbes and prevent them from infecting cells, thus “neutralizing” the microbes. IgG antibodies coat (“opsonize”) microbes and target them for phagocytosis, since phagocytes (neutrophils and macrophages) express receptors for the Fc tails of IgG. IgG and IgM activate the complement system by the classical pathway, and complement products promote phagocytosis and destruction of microbes. The production of most opsonizing and complement-fixing IgG antibodies is stimulated by TH1 helper cells, which respond to many bacteria and viruses; thus, the protective response to most bacteria and viruses is driven by TH1 cells. http://classconnection.s3.amazonaws.com/817/flashcards/995817/jpg/formation_of_secretory_iga1323598120594.jpg
 microbes and target them for phagocytosis, since phagocytes (neutrophils and macrophages) express receptors for the Fc tails of IgG. IgG and IgM activate the complement system by the classical pathway, and complement products promote phagocytosis and destruction of microbes. The production of most opsonizing and complement-fixing IgG antibodies is stimulated by TH1 helper cells, which respond to many bacteria and viruses; thus, the protective response to most bacteria and viruses is driven by TH1 cells.")
32
http://faculty.ccbcmd.edu/courses/bio141/lecguide/unit5/intro/gensteps/images/u3fg18b.jpg http://classconnection.s3.amazonaws.com/799/flashcards/1565799/png/capture1337446126520.png

33
Some antibodies serve special roles at particular anatomic sites. IgA is secreted from mucosal epithelia and neutralizes microbes in the lumens of the respiratory and gastrointestinal tracts (and other mucosal tissues). IgG is actively transported across the placenta and protects the newborn until the immune system becomes mature. IgE and eosinophils cooperate to kill parasites, mainly by release of eosinophil granule contents that are toxic to the worms. TH2 cells secrete cytokines that stimulate the production of IgE and activate eosinophils, and thus the response to helminths is orchestrated by TH2 cells. http://faculty.ccbcmd.edu/courses/bio141/lecguide/unit5/humoral/5classes/images/u3fig2t.jpg
. IgG is actively transported across the placenta and protects the newborn until the immune system becomes mature. IgE and eosinophils cooperate to kill parasites, mainly by release of eosinophil granule contents that are toxic to the worms. TH2 cells secrete cytokines that stimulate the production of IgE and activate eosinophils, and thus the response to helminths is orchestrated by TH2 cells.")
34
Most circulating IgG antibodies have half-lives of about 3 weeks. Some antibody-secreting plasma cells migrate to the bone marrow and live for years, continuing to produce low levels of antibodies. The majority of effector lymphocytes induced by an infectious pathogen die by apoptosis after the microbe is eliminated, thus returning the immune system to its basal resting state, called homeostasis. The initial activation of lymphocytes also generates long-lived memory cells, which may survive for years after the infection. Memory cells are an expanded pool of antigen-specific lymphocytes (more numerous than the naive cells specific for any antigen that are present before encounter with that antigen), and that respond faster and more effectively when re-exposed to the antigen than do naive cells. This is why the generation of memory cells is an important goal of vaccination.
, and that respond faster and more effectively when re-exposed to the antigen than do naive cells. This is why the generation of memory cells is an important goal of vaccination..")
35
http://www.nature.com/nri/journal/v3/n10/images/nri1201-i1.jpg

36
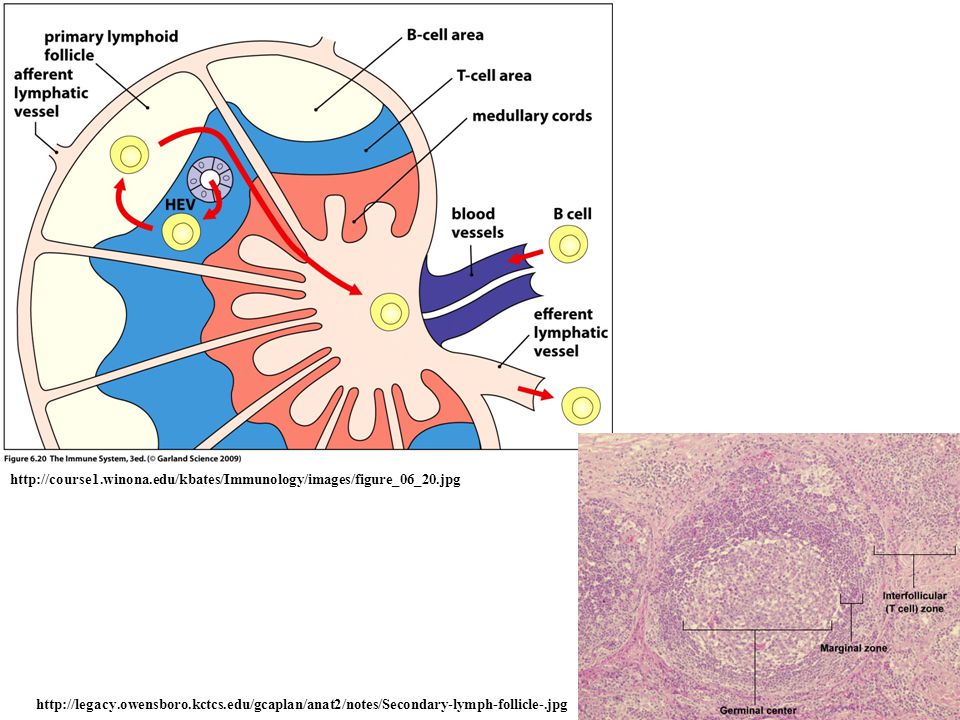
http://course1.winona.edu/kbates/Immunology/images/figure_06_20.jpg http://legacy.owensboro.kctcs.edu/gcaplan/anat2/notes/Secondary-lymph-follicle-.jpg
38
T Lymphocytes T lymphocytes develop from precursors in the thymus. Mature T cells are found in the blood, where they constitute 60% to 70% of lymphocytes, and in T-cell zones of peripheral lymphoid organs (described below). Each T cell recognizes a specific cell-bound antigen by means of an antigen-specific T-cell receptor (TCR). In approximately 95% of T cells the TCR consists of a disulfide-linked heterodimer made up of an α and a β polypeptide chain, each having a variable (antigen- binding) region and a constant region.
. Each T cell recognizes a specific cell-bound antigen by means of an antigen-specific T-cell receptor (TCR). In approximately 95% of T cells the TCR consists of a disulfide-linked heterodimer made up of an α and a β polypeptide chain, each having a variable (antigen- binding) region and a constant region..")
40
The αβ TCR recognizes peptide antigens that are displayed by major histocompatibility complex (MHC) molecules on the surfaces of antigen-presenting cells (APCs). By limiting the specificity of T cells for peptides displayed by cell surface MHC molecules, called MHC restriction, the immune system ensures that T cells see only cell-associated antigens (e.g., those derived from microbes in cells). http://life.nthu.edu.tw/~b861610/seminar/2000-4-13/tcr-mhc.JPG
.")
41
TCR diversity is generated by somatic rearrangement of the genes that encode the TCR α and β chains. All cells of the body, including lymphocyte progenitors, contain TCR genes in the germ-line configuration, which cannot be expressed as TCR proteins. During T cell development in the thymus, the TCR genes rearrange to form many different combinations that can be transcribed and translated into functional antigen receptors.

42
The enzyme in developing lymphocytes that mediates rearrangement of antigen receptor genes is the product of RAG-1 and RAG-2 (recombination activating genes); inherited defects in RAG proteins result in a failure to generate mature lymphocytes. Whereas each T cell expresses TCR molecules of one specificity, collectively, the full complement of T cells in an individual is capable of recognizing a very large number of antigens. It is important to note that unrearranged (germ-line) TCR genes are present in all non-T cells in the body, but only T cells contain rearranged TCR genes. Hence, the presence of rearranged TCR genes, which can be demonstrated by molecular analysis, is a marker of T-lineage cells. Furthermore, because each T cell and its clonal progeny have a unique DNA rearrangement (and hence a unique TCR), it is possible to distinguish polyclonal (non-neoplastic) T-cell proliferations from monoclonal (neoplastic) T-cell proliferations. Thus, analysis of antigen receptor gene rearrangements is a valuable assay for detecting lymphoid tumors.
 TCR genes are present in all non-T cells in the body, but only T cells contain rearranged TCR genes. Hence, the presence of rearranged TCR genes, which can be demonstrated by molecular analysis, is a marker of T-lineage cells. Furthermore, because each T cell and its clonal progeny have a unique DNA rearrangement (and hence a unique TCR), it is possible to distinguish polyclonal (non-neoplastic) T-cell proliferations from monoclonal (neoplastic) T-cell proliferations. Thus, analysis of antigen receptor gene rearrangements is a valuable assay for detecting lymphoid tumors..")
43
http://wenliang.myweb.uga.edu/mystudy/immunology/ScienceOfImmunol ogy/NotesImages/Topic174NotesImage5.gif

44
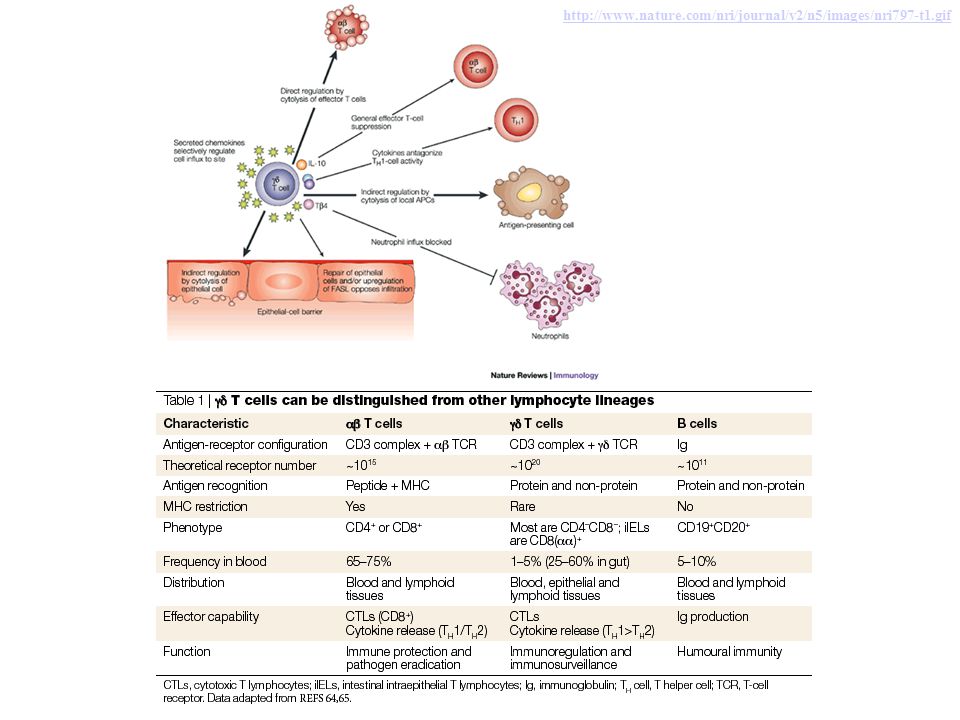
A small population of mature T cells expresses another type of TCR composed of γ and δ polypeptide chains. The γδ TCR recognizes peptides, lipids, and small molecules, without a requirement for display by MHC proteins. γδ T cells tend to aggregate at epithelial surfaces, such as the skin and mucosa of the gastrointestinal and urogenital tracts, suggesting that these cells are sentinels that protect against microbes that try to enter through epithelia. However, the functions of γδ T cells are not clearly understood. Another small subset of T cells expresses markers that are found on NK cells; these cells are called NK-T cells. NK-T cells express a very limited diversity of TCRs, and they recognize glycolipids that are displayed by the MHC-like molecule CD1. The functions of NK- T cells are also not well defined.

45
Figure 1 Structure of the TCR Tripodo, C. et al. (2009) Gamma-delta T-cell lymphomas Nat. Rev. Clin. Oncol. doi:10.1038/nrclinonc.2009.169

46
During T-cell development, CD4 − CD8 − T-cells are committed either to an αβ or γδ fate as a result of an initial β or δ TCR gene rearrangement. Cells that undergo early β chain rearrangement express a pre-TCR structure composed of a complete β chain and a pre-TCRα chain on the cell surface. Such cells switch to a CD4 + CD8 + state, rearrange the TCRα chain locus, and express a mature αβ TCR on the surface. CD4 − CD8 − T cells that successfully complete the γ gene rearrangement before the β gene rearrangement express a functional γδ TCR and remain CD4 − CD8 −. The complete α, β, γ, and δ TCR chains display strong structural homology with immunoglobulin light and heavy chains, as they consist of a variable amino-terminal region and a constant region. Abbreviations: C, constant; DN, double-negative (CD4 − CD8 − ); DP, double-positive (CD4 + CD8 + ); pTα, pre-TCRα chain; TCR, T-cell receptor; V, variable. Figure legend for previous slide
; DP, double-positive (CD4 + CD8 + ); pTα, pre-TCRα chain; TCR, T-cell receptor; V, variable. Figure legend for previous slide.")
47
http://www.nature.com/nri/journal/v2/n5/images/nri797-t1.gif
48
In addition to CD3 and ζ proteins, T cells express several other proteins that assist the TCR complex in functional responses. These include CD4, CD8, CD2, integrins, and CD28. CD4 and CD8 are expressed on two mutually exclusive subsets of αβ T cells.

49
CD4 is expressed on approximately 60% of mature CD3+ T cells, which function as cytokine-secreting helper cells that help macrophages and B lymphocytes to combat infections, whereas During antigen presentation, CD4 molecules bind to class II MHC molecules that are displaying antigen.

50
http://ars.els-cdn.com/content/image/1-s2.0-S095279151200115X-gr2.jpg

51
CD8 is expressed on about 30% of T cells, which function as cytotoxic (killer) T lymphocytes (CTLs) to destroy host cells harboring microbes. CD4 and CD8 serve as “coreceptors” in T-cell activation, so called because they work with the antigen receptor in responses to antigen.

52
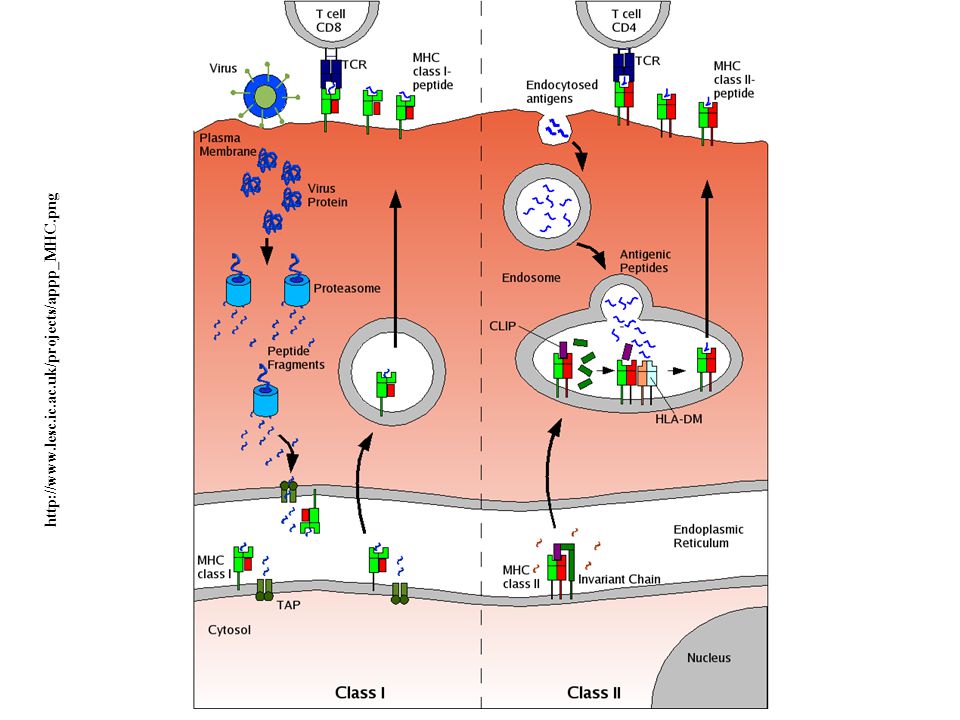
When the antigen receptor of a T cell recognizes antigen, the CD4 or CD8 co-receptor initiates signals that are necessary for activation of the T cells. Because of this requirement for co-receptors, CD4+ helper T cells can recognize and respond to antigen displayed only by class II MHC molecules, whereas CD8+ cytotoxic T cells recognize cell-bound antigens only in association with class I MHC molecules.

54
Dendritic Cells There are two types of cells with dendritic morphology that are functionally quite different. Both have numerous fine cytoplasmic processes that resemble dendrites, from which they derive their name.

55
http://www.nobelprize.org/educational/medicine/immunity//images/detail/agp.gif Interdigitating dendritic cells, or just dendritic cells. These cells are the most important antigen-presenting cells (APCs) for initiating primary T-cell responses against protein antigens. Several features of dendritic cells account for their key role in antigen presentation.
 for initiating primary T-cell responses against protein antigens. Several features of dendritic cells account for their key role in antigen presentation..")
56
http://upload.wikimedia.org/wikipedia/commons/2/2e/Dendritic_cells.jpg First, these cells are located at the right place to capture antigens—under epithelia, the common site of entry of microbes and foreign antigens, and in the interstitia of all tissues, where antigens may be produced. Immature dendritic cells within the epidermis are called Langerhans cells.

57
http://www.nature.com/nri/journal/v1/n2/images/nri1101-135a-f5.gif Second, dendritic cells express many receptors for capturing and responding to microbes (and other antigens), including TLRs and mannose receptors.
, including TLRs and mannose receptors.")
58
Third, in response to microbes, dendritic cells are recruited to the T- cell zones of lymphoid organs, where they are ideally located to present antigens to T cells. Fourth, dendritic cells express high levels of the molecules needed for presenting antigens to and activating CD4+ T cells.

59
Follicular dendritic cell These cells bear Fc receptors for IgG and receptors for C3b and can trap antigen bound to antibodies or complement proteins. Such cells play a role in humoral immune responses by presenting antigens to B cells and selecting the B cells that have the highest affinity for the antigen, thus improving the quality of the antibody produced. http://www.nature.com/nri/journal/v3/n9/images/nri1179-f3.gif http://www.nature.com/nri/journal/v3/n9/images/nri1179-f4.gif http://www.fitzgerald-fii.com/media/catalog/product/cache/1/small_image/9df78eab33525d08d6e5fb8d27136e95/f/o/follicular-dendritic-cells-antibody-10r-m164a_IHC.jpg.jpg

60
MHC is HLA HLA and Disease Association Disease HLA Allele Risk (%) Ankylosing spondylitisB2790– 100 Postgonococcal arthritisB2714 Acute anterior uveitisB2714 Rheumatoid arthritisDR44 Chronic active hepatitisDR313 Primary Sjogren syndrome DR39 Type 1 diabetesDR35 DR46 DR3/DR420
 Ankylosing spondylitisB2790– 100 Postgonococcal arthritisB2714 Acute anterior uveitisB2714 Rheumatoid arthritisDR44 Chronic active hepatitisDR313 Primary Sjogren syndrome DR39 Type 1 diabetesDR35 DR46 DR3/DR420")
61
APCs All nucleated cells

62
http://www.lesc.ic.ac.uk/projects/appp_MHC.png
63
http://www.jimmunol.org/content/182/6/3335/F3.large.jpg

Similar presentations





 or antibody mediated immunity: The total immunological.>")






 Features of immune.>")

>")




 - any substance, which is recognized by the mature immune system of a given organism.>")



