Download presentation
Presentation is loading. Please wait.

1
Lecture 22 announcements: Course evaluation reminder, before Dec 7 Under Lecture Syllabus find Topics per lecture and keywords Under Essays source papers find Membracid helmet function Under Essays special topics find Tentorium

3
Morphological diversity in treehopper pronota/helmets
B Prud’homme et al. Nature 473, (2011) doi: /nature09977
 doi: /nature")
4
Sculpture by Alfred Keller Berlin Nature Museum
Is this structure homologous with the ‘lost’ prothoracic wings of Insecta?

5
Prud’home, B. et al Body plan innovation in treehoppers through the evolution of an extra wing-like appendage. Nature 473: 83-86 From Wikki: membracids are “… known for their enlarged and ornate pronotum, which most often resembles thorns, apparently to aid camouflage… The specialised pronotum (or helmet) may not be simply an expansion of the prothoracic sclerite, but a fused pair of dorsal appendages of the first thoracic segment. These [fused appendages] may be serial homologues of insect wings, which are dorsal appendages of the second and/or third thoracic segments. Evidence for this theory includes the development of the helmet, which arises as a pair of appendages attached to each side of the dorsal prothorax by an articulation with muscles and a flexible membrane that allow it to be mobile. Also, the same [Hox] genes are involved in development of the helmet and the wings.” Read my essay online, written before I read a paper by Yoshizawa (2012) quarrelling with this hypothesis.
 may not be simply an expansion of the prothoracic sclerite, but a fused pair of dorsal appendages of the first thoracic segment. These [fused appendages] may be serial homologues of insect wings, which are dorsal appendages of the second and/or third thoracic segments. Evidence for this theory includes the development of the helmet, which arises as a pair of appendages attached to each side of the dorsal prothorax by an articulation with muscles and a flexible membrane that allow it to be mobile. Also, the same [Hox] genes are involved in development of the helmet and the wings. Read my essay online, written before I read a paper by Yoshizawa (2012) quarrelling with this hypothesis.")
6
[Serial homology: structures are considered serial homologues if they represent corresponding parts of organisms built according to the same body plan. For example, the notopodia of Nereis are homologues. The body plan of the insect thorax is three segments in series, each segment with a pair of legs ventrally and a pair of wings dorsally. Treehoppers, alone among modern insects, have evolved to ‘turn back on’ genes that made the beginnings of wings, genes shut down in all other modern insects. Information used long long ago in development of the prothoracic metamere of insects and normally in modern insects turned off by Hox genes, is somehow allowed to proceed. In membracids it begins to form a pair of articulated structures [wing primordia] but these do not differentiate to wings, they just begin. Other selection later acted on genes subsequently affecting these primordia to give features of the helmet. Yoshizawa, K The treehopper’s helmet is not homologous with wings (Hemiptera: Membracidae). Systematic Entomology 37: 2-6. “Prud’homme et al. have misinterpreted a part of T2 as T1….all the nymphal characteristics mentioned by Prud’homme et al. (2011) are interpreted incorrectly.”
. Systematic Entomology 37: 2-6. Prud’homme et al. have misinterpreted a part of T2 as T1….all the nymphal characteristics mentioned by Prud’homme et al. (2011) are interpreted incorrectly.")
7
Critically one looks for structures developing in the region Prud’homme et al. identify as a wing hinge that are homologues of the axillary sclerites, pleural wing process etc. “Evidence for the ‘joint between the helmet and T1 tergum is crucial…” A membranous region of cuticle that Prud’homme say serves as a hinge is misinterpreted: it is actually “the intersegmental connection between T1 and T2…” This region is not homologous with the wing joint membranes (hinges) of T2 and T3.
 of T2 and T3.")
8
Sensory processing: transduction of pressure into neuron depolarizations
Organs called ears are mechanical transducers and their essence is a tympanum or eardrum which tracks the pressure [or displacement] changes that associate with sound travelling through [the fluid] air. Insects have trachea, air-filled tubes coursing through their body to convey gases. The tracheal system plays an important part in the workings of ears. To ‘collect’ pressure changes requires a thin membrane of cuticle and this must be free to move readily – it cannot be damped by body fluids. So we have a tympanum backed by a tracheal air sac. The ear of insects is typically a pressure difference ear in that there are two routes to the eardrum: internal and external. The movement of the eardrum is thus a compounded effect of two path-lengths which may be different. The moving membrane has special cells linked to it that perform the actual transduction: converting pressure forces into depolarizing membranes (the ‘information currency’ of the nervous system.

9
What is an ear and how does a locust discriminate sound frequencies?
Ears are typically bilateral part of the bilateral symmetry of animals, a right and a left. In the locust these are pressure difference ears; they are each situated within a recess in the first abdominal segment and sound has access to the rear of both ears (pressure difference). The plane of the tympanum is angled to face backward slightly. The auditory ganglion of each ear is visible through the transparent tympanum, its nerve running anteromedially to join the metathoracic ganglion. Also visible on and through the tympanum are dark brown chitinous structures (e.g., pyriform vesicle, folded body) that lie on top of the tympanum.
. The plane of the tympanum is angled to face backward slightly. The auditory ganglion of each ear is visible through the transparent tympanum, its nerve running anteromedially to join the metathoracic ganglion. Also visible on and through the tympanum are dark brown chitinous structures (e.g., pyriform vesicle, folded body) that lie on top of the tympanum.")
10
At one time it was disputed whether insects could discriminate frequency (indeed whether they even had hearing capacity) [insect baloonists experiments]. First the anatomy using an anchient note but drawing in chalk. Easier to follow when one sees a diagram made in front of you.
![At one time it was disputed whether insects could discriminate frequency (indeed whether they even had hearing capacity) [insect baloonists experiments].](http://slideplayer.com/slide/3282023/11/images/10/At+one+time+it+was+disputed+whether+insects+could+discriminate+frequency+%28indeed+whether+they+even+had+hearing+capacity%29+%5Binsect+baloonists+experiments%5D..jpg "First the anatomy using an anchient note but drawing in chalk. Easier to follow when one sees a diagram made in front of you.")
11
The tympanum (ear drum) is a very-much thinned region of the cuticle with a ganglion sitting more or less in the middle. Behind the tympanum, applied overtop of ganglion and acoustic nerve is a tracheal sac. Backing the membrane with air is an important adaptation: if the tympanum were backed by haemolymph of the circulatory system the tympanum’s movements would be significantly damped by the blood and it would not respond with sensitivity to the ariborne sound.

12
Chordotonal sensilla occur in the ganglion, 60 to 80 in four groupings; each sensillum involves several cell types. The sensillum transduces the mechanical movements of the pyriform vesicle or other cuticular eardrum parts. When the modified dendritic region surrounded by the scolopale cell is mechanically stimulated (by sound and resulting eardrum movement) the axon develops an action potential and the neurosensory cell, by firings, sends information to the CNS. The cell’s position on the eardrum, its mechanical linkage, and the behaviour of the eardrum itself, codes for particular frequencies.
 the axon develops an action potential and the neurosensory cell, by firings, sends information to the CNS. The cell’s position on the eardrum, its mechanical linkage, and the behaviour of the eardrum itself, codes for particular frequencies..")
13
fusiform body pyriform vesicle
Fusiform body (fb) and pyriform vesicle (pv) and frequency discrimination. At 3-kHz sound input the whole ganglion follows the motion of the pyriform vesicle (pv); so both ends move in phase. But at the higher frequency of 10 kHz the relative motions of ganglion (K1) vs pv are quite different and the strand of nervous tissue (fb) is shaken and jolted, leading to many firings of the chordotonal neurosensory cells within. fusiform body pyriform vesicle Stephen R.O., Bennet-Clark H.C The anatomical and mechanical basis of stimulation and frequency analysis in the locust ear. J. exp. Biol. 99: Stephen R.O., Bennet-Clark H.C The anatomical and mechanical basis of stimulation and frequency analysis in the locust ear. J. exp. Biol. 99:
 and pyriform vesicle (pv) and frequency discrimination. At 3-kHz sound input the whole ganglion follows the motion of the pyriform vesicle (pv); so both ends move in phase. But at the higher frequency of 10 kHz the relative motions of ganglion (K1) vs pv are quite different and the strand of nervous tissue (fb) is shaken and jolted, leading to many firings of the chordotonal neurosensory cells within. fusiform body. pyriform. vesicle. Stephen R.O., Bennet-Clark H.C The anatomical and mechanical basis of stimulation and frequency analysis in the locust ear. J. exp. Biol. 99: Stephen R.O., Bennet-Clark H.C The anatomical and mechanical basis of stimulation and frequency analysis in the locust ear. J. exp. Biol. 99:")
14
James F. C. Windmill, Martin C. Gopfert and Daniel Robert 2004
James F.C. Windmill, Martin C. Gopfert and Daniel Robert Tympanal travelling waves in migratory locusts Journal of experimental Biology 208: Scanning laser vibrometry used to investigate the movements of the eardrum when stimulated by different frequencies. James F.C. Windmill, Martin C. Gopfert and Daniel Robert Tympanal travelling waves in migratory locusts Journal of experimental Biology 208:

15
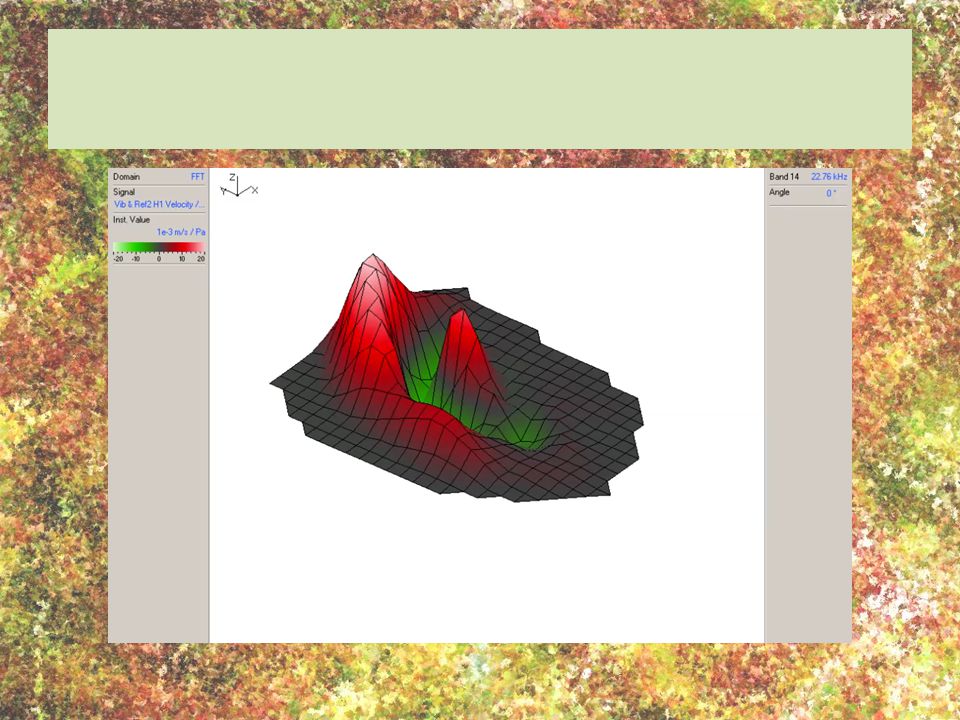
Frequency analysis in the locust involves a “travelling wave that funnels mechanical energy to specific tympanal locations, where distinct mechanoreceptor neurones project”. “For each frequency the tympanal deflections do not stay in position, but travel across the tympanum from posterior to anterior... At 3.3 kHz the wave travels across the thin membrane, moving towards a focus point located at the folded body...” Travelling waves vs standing waves eardrum movement when subjected to four different frequencies; scanning laser videos show the complex movement of different regions; profiles: red is outward movement of the tympanum and green is inward movement

18
What can one tell about function from the morphology of a neuron?
Giant neurons occur in many animals, e.g., Annelids, Arthropods, Molluscs. They are adaptive in emergency escape: sabellid worms use giant neurons to retract their ‘lophophore’ into the safety of their tubes when attacked. There is no ‘consulting’ with the brain: speed is essential. These cells were used by early physiologists trying to understand depolarization of the nerve cell membrane. In 1936 J.Z. Young discovered that certain long structures present in squids (previously thought to be blood vessels) were actually single unusually large neurons. They had axons of unusually large diameter: up to 1 mm. A typical axon is about 40 microns in diameter: these giants are about 700 microns. They run through the length of the body. Kenneth Roeder and cockroach experiments.
 were actually single unusually large neurons. They had axons of unusually large diameter: up to 1 mm. A typical axon is about 40 microns in diameter: these giants are about 700 microns. They run through the length of the body. Kenneth Roeder and cockroach experiments.")
19
Stellate ganglion in squid mantle
Stellate ganglion in squid mantle. The stellate nerve contains a giant axon. This large-diametered motor nerve depolarizes very rapidly, ensuring nearly synchronous activation of mantle muscles in a jet-propelled escape. Varying thickness promotes different speeds of conductivity: longer are thicker and go faster so everything happens at once. Pictures taken from a website illustrating the dissecting out of a nervous preparation of the squid giant synapse (USCRC IBRO)
")
21
Interneurons and shape specificity (From J. Insect Physiology, George Boyan, 1984) Interneurons that receive auditory input: note topographical similarity of omegas of cricket Gryllus and katydid Tettigonia, reflecting binaural comparison. Neurons can be identified across taxa based upon their morphology, e.g., making crossbody comparisons or as in the case of AN1 conveying activity from the prothoracic ganglion to the brain. efferent, afferent Analogous to wiring, a nerve or a neuron says something about its function by where it starts and where it winds up: a motor message going from the CNS to a certain muscle (efferent), a sensory message returning from an eardrum to the CNS (afferent).
 Interneurons that receive auditory input: note topographical similarity of omegas of cricket Gryllus and katydid Tettigonia, reflecting binaural comparison. Neurons can be identified across taxa based upon their morphology, e.g., making crossbody comparisons or as in the case of AN1 conveying activity from the prothoracic ganglion to the brain. efferent, afferent. Analogous to wiring, a nerve or a neuron says something about its function by where it starts and where it winds up: a motor message going from the CNS to a certain muscle (efferent), a sensory message returning from an eardrum to the CNS (afferent).")
22
Interneurons are shaped adaptively: the story of the omega neuron of orthopteran insects
The adaptiveness of this neuron’s shape relates to bilateral perception of sound by right and left ears, located on the front legs of crickets and katydids. sound localization mechanism and contralateral inhibition. There are two overlapped mirror-image interneurons; each feeding back upon the other. Gerry Pollock

23
Scientific American Huber & Thorson
Pressure difference ear with 4 different inputs phase shifter omega interneuron is in the prothoracic ganglion

24
Adaptiveness of omega shape of the neuron of a cricket
Omega interneuron: its name suggested by the shape, lies within the prothoracic ganglion of a cricket or katydid. It receives input from the acoustic nerve running back up the foreleg from the ears. Parts of a neuron: cell body (soma), dendritic arborization, axon. Omega neuron is here filled with green dye via the electrode that once monitored its firing activity; omega neurons occur in the prothoracic ganglion of a cricket as an overlain mirror-image pair; the firing of each feeds back upon and inhibits the activity of the other. Thus a slight difference in perception to one side is enhanced, supporting better localization. Gerry Pollack, Montreal Contralateral inhibition
, dendritic arborization, axon. Omega neuron is here filled with green dye via the electrode that once monitored its firing activity; omega neurons occur in the prothoracic ganglion of a cricket as an overlain mirror-image pair; the firing of each feeds back upon and inhibits the activity of the other. Thus a slight difference in perception to one side is enhanced, supporting better localization. Gerry Pollack, Montreal. Contralateral inhibition.")
25
SOUND LOCALIZATION: DETERMINING DIRECTION AND DISTANCE TO A SOUND SOURCE. How does a too-small female field cricket find her mate by phonotaxis using his too-long sound wave? We determine direction to sound sources by comparing IIDs: interaural intensity differences. These differences arise in a right and left ear because of different path lengths (one ear is closer to the sound than the other) and they arise because of sound diffraction (bending) about the body. Small bodies relative to sound wavelength have minimal diffraction. Large bodies make effective obstacles and create a ‘sound shadow’. Human ears are separated by ~21 cm. This distance is about the wavelength of a 800 Hz sound. For this wavelength the (diffraction) cast by the human head gives an intensity difference of about 8 dB. For higher frequencies, e.g., a tone of 10 kHz, this effect becomes more pronounced, e.g., a 20 dB drop in intensity at the farther ear for 10 kHz. Humans make use of IIDs, turning their head to equalize sound levels and by doing so facing in the direction of the source of a sound. Assigned reading: Michelsen, A The tuned cricket. New Physiol. Sci. 13: Michelsen, A. & Lohe, G Tuned directionality in cricket ears. Nature 375: 639- Michelsen A. et al Physics of directional hearing in the cricket Gryllus bimaculatus J. comp. Physiol. A 175:

26
* Explain phase. Cricket directionality is achieved because the ears are pressure gradient ears, i.e., sound has access to both front and rear of the eardrum, and also because in crickets this back access involves cross-body transfer of sound in a large prothoracic transverse trachea. The activity of the eardrum is a resultant of pressure changes both outside and inside. And at any given orientation of the female cricket, the net pressure at each ear is governed by phase produced by path-length differences. The physiologists refer to IIDs: interaural intensity differences. The different path lengths to the rear of the eardrum create phase differences which activate right and left eardrums to differing extent. Phase changes with orientation.

27
Because a cricket’s body is small relative to the song sound wavelengths it broadcasts, it cannot create useful interaural intensity differences IIDs by diffraction. Cricket call wavelength is about 70 mm; the distance between the leg-situated ears with the legs in walking position, is no more than 10 mm. The body as an obstacle causes no significant drop in intensity to a farther ear. The female cricket can turn 360º in relation to a distant calling male and IIDs remain nearly the same. Yet in fact female crickets localize male calls well, at night, at a distance and can readily walk toward that sound source. How do they do it?

28
* *ipsilateral same side as source; contralateral opposite side Imagine sound from a male’s call reaching the front of the near eardrum. It also arrives at the inner surface of this same eardrum via 3 other routes : two prothoracic spiracles on the thorax (IS & CS: ipsilateral* spiracle and contralateral spiracle) and the contralateral tympanum. The path lengths of the three routes change as the female turns. Thus the sound pressures on the back of the eardrum will change with changing phase relative to those on the outside. So right and left eardrums show different activity as a function of body direction relative to the source. Sound reaching the back of the eardrum later than sound reaching the front is shifted in time, i.e., its phase has changed.
 and the contralateral tympanum. The path lengths of the three routes change as the female turns. Thus the sound pressures on the back of the eardrum will change with changing phase relative to those on the outside. So right and left eardrums show different activity as a function of body direction relative to the source. Sound reaching the back of the eardrum later than sound reaching the front is shifted in time, i.e., its phase has changed.")
29
Michelsen broadcast 4.5 kHz to female crickets in an arena, moving a speaker to 12 different surrounding positions and monitoring eardrum activity with a laser vibrometer. The dotted line P is a vector (magnitude and direction) for the driving force (pressure) at the eardrum for these 12 sound-source directions. The resulting directional pattern for the right ear is asymmetrical, i.e., as the phase angle changes there are side-to-side differences in eardrum activity which can be transduced and compared by the omega neurons of the prothoracic ganglion. Now the cricket has IIDs and a way of localizing the sound source in the absence of effective diffraction.
 for the driving force (pressure) at the eardrum for these 12 sound-source directions. The resulting directional pattern for the right ear is asymmetrical, i.e., as the phase angle changes there are side-to-side differences in eardrum activity which can be transduced and compared by the omega neurons of the prothoracic ganglion. Now the cricket has IIDs and a way of localizing the sound source in the absence of effective diffraction..")
Similar presentations


















>")



