Download presentation
Presentation is loading. Please wait.

1
Bio 325 Lecture 15 ‘Sudden appendages’: lunging and jumping Roberts T.J. & Azizi E. 2011. Flexible mechanisms: the diverse roles of biological springs in vertebrate movement. J. exp. Biol. 214: 353-361. “Springs deform when a force is applied and recoil to their resting shape when the force is released. Materials [resilin, abductin are just extremes of rubbery flexible skeletal materials -- chitin and collagen also store energy even when relatively stiff] can act like springs when loaded in tension, like a rubber band, or in compression, like a rubber ball. Both kinds of loading are important in nature. When springy materials deform, they store energy in the form of elastic strain energy, and when they recoil this energy is released. The amount of energy stored depends on the material stiffness and the deformation.” “Elastic mechanisms can act as power amplifiers, by storing muscle work slowly and releasing it rapidly.” This is the flea using resilin as described last lecture.” The jump of the flea involves a kind of spring enabling power amplification, widespread in animal locomotion, both invertebrates and vertebrates.

2
Sources re sudden appendages *Assigned reading *Heitler, W.J. 1974. The locust jump, specialisations of the metathoracic femoral-tibial joint. Journal of comparative Physiology 89: 93-104. *Bayley, T.G. 2012. A buckling region in locust hindlegs contains resilin and absorbs energy when jumping or kicking goes wrong. J. exp. Biol. 215: 1151-1161. *Patek, S.N. et al. 2011. From bouncy legs to poisoned arrows: elastic movements in invertebrates. J. exp. Biol. 214: 1973-1980. * Rothschild, M. et al. 1973. The flying leap of the flea. Scientific American 222: 92-101 Gordon, J.E. 1978. Structures or Why Things Don’t Fall Down. Penguin. Sutton, G.P. & Burrows, M. 2011. Biomechanics of jumping in the flea. J. exp. Biol. 214: 836-847.

3
Romaleinae Eumastacinae Theme of the course applied to the metathoracic leg of Orthoptera. This colourful insect (left) is a eumasticid grasshopper from Colombia.
 is a eumasticid grasshopper from Colombia..")
4
Saltatorial: animal modified for leaping and jumping. Forelimbs small, hindlimbs enlarged tracks in snow

5
isometric contraction and resilin the basis of high-power leaping in fleas figure is from Rothschild’s Scientific American article isometric means antagonistic muscles generate force without changing length

6
Sutton G.P., Burrows M. 2011. Biomechanics of jumping in the flea. J. of exp. Biol. 214: 836-847. In this paper the authors present two hypotheses of how the flea jump works: 1) Rothschild Hypothesis ‘trochanters driven into ground’ 2) Bennet-Clark Hypothesis ‘overall extension of leg speeded up’. They decide in favour of the latter hypothesis: the trochanters do not touch the ground, rather the “expansion of the spring applied a torque about the coxotrochanteral joint that (is) carried through the femur and tibia and finally resulted in a force applied to the ground by the hind tibia and tarsus. In other words the whole chain of leg segments extends with enhanced speed derived from the resilin. Driving down the trochanters into the ground has some obvious arguments against: animal would be propelled vertically and could have trouble jumping and making horizontal distance (of course to reach a passing dog vertical might be rather good).
 Rothschild Hypothesis ‘trochanters driven into ground’ 2) Bennet-Clark Hypothesis ‘overall extension of leg speeded up’. They decide in favour of the latter hypothesis: the trochanters do not touch the ground, rather the expansion of the spring applied a torque about the coxotrochanteral joint that (is) carried through the femur and tibia and finally resulted in a force applied to the ground by the hind tibia and tarsus. In other words the whole chain of leg segments extends with enhanced speed derived from the resilin. Driving down the trochanters into the ground has some obvious arguments against: animal would be propelled vertically and could have trouble jumping and making horizontal distance (of course to reach a passing dog vertical might be rather good)..")
8
from Wikkipedia Locusta migratoria in a prejump crouch. Heitler W.J. 1974. The locust jump. J. comp. Physiol. 89: 93-104 Orthoptera Species File

9
Heitler W.J. 1974. The locust jump. Journal of comparative Physiology 89: 93-104. Well-written and clear; read it in detail to understand the paradox (see below). Heitler starts with behaviour: what the structure does: the mechanics of the jump in increasing detail. There is passing reference to the adaptive context. “The locust jumps to escape from danger, to launch itself for flight, or simply to achieve a more rapid form of locomotion than walking. Prior to a jump the locust assumes a crouched position, with the metathoracic tibiae flexed, and it may maintain this for some seconds until it either jumps or relaxes. The jump is achieved through a rapid extension of both the metathoracic tibiae... [through a femorotibial joint “excursion of about 150 degrees”].
. Heitler starts with behaviour: what the structure does: the mechanics of the jump in increasing detail. There is passing reference to the adaptive context. The locust jumps to escape from danger, to launch itself for flight, or simply to achieve a more rapid form of locomotion than walking. Prior to a jump the locust assumes a crouched position, with the metathoracic tibiae flexed, and it may maintain this for some seconds until it either jumps or relaxes. The jump is achieved through a rapid extension of both the metathoracic tibiae... [through a femorotibial joint excursion of about 150 degrees ]..")
11
Flexion and extension: during a jump or kick the joint angle goes from 0 to 150 º Dicondylic joint: condyles [= pivot pegs] are part of femur, the sockets part of tibia. Extensor of tibia, flexor of tibia are antagonists, yet their muscles pull in nearly the same direction. The femorotibial (dicondylic) joint is the axis about which the tibia pivots relative to the femur. The extensor works as a 1 st class lever; the flexor is 3 rd class. Notice that the lever arm is bent in a distinctive shape which affects force direction (see below). The angles of ‘force in’ change as the tibia moves from completely flexed to maximally extended (see below). Pocket and lump = flexor apodeme lock. Anatomy and leverage of locust metathoracic leg femorotibial joint
![Flexion and extension: during a jump or kick the joint angle goes from 0 to 150 º Dicondylic joint: condyles [= pivot pegs] are part of femur, the sockets part of tibia.](http://images.slideplayer.com/10/2760534/slides/slide_11.jpg "Extensor of tibia, flexor of tibia are antagonists, yet their muscles pull in nearly the same direction. The femorotibial (dicondylic) joint is the axis about which the tibia pivots relative to the femur. The extensor works as a 1 st class lever; the flexor is 3 rd class. Notice that the lever arm is bent in a distinctive shape which affects force direction (see below). The angles of ‘force in’ change as the tibia moves from completely flexed to maximally extended (see below). Pocket and lump = flexor apodeme lock. Anatomy and leverage of locust metathoracic leg femorotibial joint.")
12
Geometry/anatomy of the joint see Heitler’s Fig. 1 a Fully flexed joint, lock engaged: bifurcate pocket of apodeme of flexor sits astride the lump. Note apodeme of extensor and two accessory muscles. b Lock is disengaged and joint extended midway; flexor apodeme is now readily visible and rides pulley- like on top of the lump. c The two condyles of the dicondylic joint seen dorsally along with the lump.

13
Paradox Both muscles work with a poor mechanical advantage due to constraints of body shape* but a good speed-distance advantage – its good to have speed and distance working for you when trying to jump. “Myograms show that there is co-activation of the extensor and flexor muscles during the pre-jump crouch...” “ Complete extension of the tibia takes some 20 ms... to develop peak power...in the jump the extensor muscle must first build up tension isometrically.” The two antagonists simultaneously contract, but there is no movement at the joint. “The extensor muscle is much larger and occupies the greater part of the femoral volume.” Its pinnate [angled like a feather] fibres are many more and short; an arrangement which enables the muscle to develop a very large force at its tendon [apodeme], though moving through a shorter distance. The flexor muscle, by contrast, is composed of long thin parallel fibres, and is of comparatively small cross-sectional area. “This weak muscle must hold the tibia flexed against the full force of the powerful extensor muscle.” How is this achieved during isometric contraction? The answer: special adaptations of leverage at the joint and a flexor apodeme lock. *For the force/effort arm of the tibia to have greater length relative to its load arm giving better advantage, we would have to change the shape of the femur’s distal end drastically.

14
We need to think about moments of force in analysing the movements at the joint. Here is a diagram based upon Heitler’s Fig. 2 c. “The thick blue and purple lines “represent a mechanical analogue of the joint structure”.

15
Go back to the paradox: how is it that during isometric contraction the smaller, weaker muscle is able to match the effort of the larger? The force advantage of the flexor muscle is different at different angles of flexion of the femorotibial joint. Part of the answer to the above question is that when the angle of flexion is less than 5 degrees (top) the mechanical advantage of the flexor muscle is superior to that of the extensor.
 the mechanical advantage of the flexor muscle is superior to that of the extensor..")
16
With the joint angle at 5º (top), the apodeme of the flexor makes an angle with the effort arm of the lever of almost 90º; this is because it rides up over the 'lump'. The lump functions as a pulley: (a pulley changes force direction); it changes the ‘force-in’ direction of the flexor apodeme making it nearly 90 º; by contrast the stronger extensor, exerting more force, has a force-in direction at a very poor angle of 6 º. So the moments of force for the two antagonists can be equal.
; it changes the ‘force-in’ direction of the flexor apodeme making it nearly 90 º; by contrast the stronger extensor, exerting more force, has a force-in direction at a very poor angle of 6 º. So the moments of force for the two antagonists can be equal..")
17
Locust gets extra force into its leap by isometric contraction. This distorts the exoskeleton in the neighbourhood of the joint and so stores elastic energy in the leg exoskeleton (semilunar processes, see below) that will later be released during the jump. This stored energy, available much more quickly than it was stored adds power. It contributes to the forces the leg exerts against the ground. The pocket is pulled over the lump during the early stages of the flexion and so the distortion can be retained even if the antagonists relax, i.e., the leg can be ‘cocked’.
 that will later be released during the jump. This stored energy, available much more quickly than it was stored adds power. It contributes to the forces the leg exerts against the ground. The pocket is pulled over the lump during the early stages of the flexion and so the distortion can be retained even if the antagonists relax, i.e., the leg can be ‘cocked’..")
18
The effort arm (the distance between the point of insertion of the apodeme on the tibia and the axis of rotation) is quite short for the extensor; much longer for the flexor. So the moments of force can be balanced at 5º. To keep the two leg muscles in isometric contraction the moment of force of the weaker muscle has been made equal to the moment of force of the stronger

19
Burrows M., Sutton G.P. 2012. Locusts use a composite of resilin and hard cuticle as an energy store for jumping and kicking. J. exp. Biol. 215: 3501-3512. Burrows & Sutton have explained where the energy of isometric contraction is stored. It goes into paired semilunar processes of the femur, located at the sides of its distal extremity, lateral to where the condyles protrude into the sockets of the tibia. semilunar process

20
Burrows & Sutton 2012 “The inside surface of a semi-lunar process consists of a layer of resilin, particularly thick along an inwardly pointing ridge and tightly bonded to the external, black cuticle.” There is (shown by imaging [movie]) distortion/bending in all three dimensions during the isometric contraction.
![Burrows & Sutton 2012 The inside surface of a semi-lunar process consists of a layer of resilin, particularly thick along an inwardly pointing ridge and tightly bonded to the external, black cuticle. There is (shown by imaging [movie]) distortion/bending in all three dimensions during the isometric contraction.](http://images.slideplayer.com/10/2760534/slides/slide_20.jpg "Burrows & Sutton 2012 The inside surface of a semi-lunar process consists of a layer of resilin, particularly thick along an inwardly pointing ridge and tightly bonded to the external, black cuticle. There is (shown by imaging [movie]) distortion/bending in all three dimensions during the isometric contraction.")
21
“Externally visible resilin was compressed and wrinkled as a semi-lunar process was bent. It then sprung back to restore the semi-lunar process to its original shape. “It is suggested that composite storage devices that combine the elastic properties of resilin with the stiffness of hard cuticle allow energy to be stored by bending hard cuticle over only a small distance and without fracturing. In this way all the stored energy is returned and the natural shape of the femur is restored rapidly so that a jump or kick can be repeated.”

23
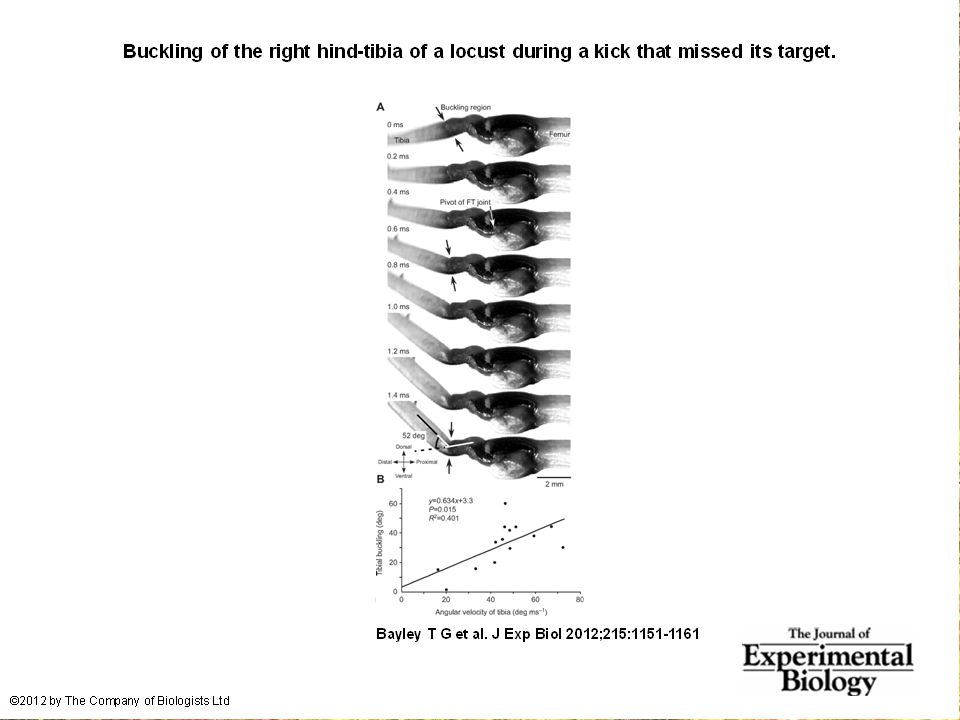
Bayley T.G., Sutton G.P., Burrows M. 2012. A buckling region in locust hindlegs contains resilin and absorbs energy when jumping or kicking goes wrong. J. exp. Biol. 215: 1151-1161. See also: JEB highlight by Kathryn Knight in same issue: Buckling zone protects locust legs Energy of a kick that misses its target (or a foot that slips on the substrate) is dissipated by a specialized proximal region of the tibia. There is resilin in this region, revealed as a band that fluoresces blue under UV illumination (with appropriate filters to confirm identity). There are also special campaniform sensilla (proprioceptors, mechanoreceptors) that monitor the buckling. “The features of the buckling region show that it can act as a shock absorber as proposed previously [by Heitler] when jumping and kicking movements go wrong.”
 is dissipated by a specialized proximal region of the tibia. There is resilin in this region, revealed as a band that fluoresces blue under UV illumination (with appropriate filters to confirm identity). There are also special campaniform sensilla (proprioceptors, mechanoreceptors) that monitor the buckling. The features of the buckling region show that it can act as a shock absorber as proposed previously [by Heitler] when jumping and kicking movements go wrong. .")
24
A bow should never be ‘shot’ without an arrow (Gordon 1976, p.92): this is because there is no way of getting rid of stored strain energy. It is possible to shatter a bow in this way. The strain energy stored in the bent bow can no longer be dissipated in the kinetic energy of the arrow and is used to make cracks in the substance of the bow. A bow makes a nice example of tension and compression surfaces in a ‘beam’ (Vogel 1988, p. 202). Bending an object of thickness has the effect of creating a gradient of tension on the outside of the curve and one of compression on the inside. In the middle there will be a neutral plane where there is no stress in either tension or compression (but not in shear). From this middle plane toward both surfaces stresses increase. This means that central regions of structures contribute less strength; it is the reason why bones can be nearly as strong when hollow. Why are the long-bones of the vertebrate limb hollow?
. Bending an object of thickness has the effect of creating a gradient of tension on the outside of the curve and one of compression on the inside. In the middle there will be a neutral plane where there is no stress in either tension or compression (but not in shear). From this middle plane toward both surfaces stresses increase. This means that central regions of structures contribute less strength; it is the reason why bones can be nearly as strong when hollow. Why are the long-bones of the vertebrate limb hollow .")
25
Special shock aborbers for kicks gone astray Gordon’s comments about bows needing arrows apply readily to locust kicks that go wrong. From the abstract of Bayley et al. 2012. “If a hindleg of a locust slips during jumping, or misses its target during kicking, energy generated by the two extensor tibiae muscles is no longer expended in raising the body [jumping] or striking a target.” Bayley found a special region of the proximal hind tibia that is adapted to buckle under these conditions through the presence of a special rubbery cuticular protein, resilin. As we now analyse in some detail the jumping of several insects, resilin will be seen to play an important role.

Similar presentations