Download presentation
Presentation is loading. Please wait.

1
Deconstruction by Enzymes 1: Cellulase 2007 Oct. 19 Speaker: Dr. Po-Huang Liang

2
CONTENT INTRODUCTION FUNDAMENTALS -Structure and Composition of Cellulosic Biomass -Cellulolytic Organisms -Cellulase Enzyme Systemes -Regulation of Cellulase Production APPLICATION OF CELLULASES -Recombinant Cellulolytic Strategy -Methodology for Studying Cellulase Properties CLOSING COMMENT

3
INTRODUCTION Need for Alternative Energy Source With the hike in oil price around the world in the 1970s and the realization that the world’s oil supply is finite, the quest for alternative fuels began in 1975. The amount of solar energy received at the earth’s surface far exceeds the amount of present human usage 2.5x10 21 Btu/year >> 2.0x10 17 Btu/year The amount of energy from sun is stored as carbon via photosynthesis, which results in production of plant biomass having cellulose as the major component

4
Strategy of converting cellulose into fuel Glucose Biofuel Biomass Cellulase

5
FUNDAMENTALS

6
Structure and Composition of Cellulosic Biomass In plants, the cell wall is constructed primarily from a carbohydrate polymer called cellulose, and the cell wall can therefore also function as a carbohydrate store for the cell. genomics.energy.gov/gallery/gtl/originals/420.jpg

7
Plant cell wall structure Plants form two types of cell wall that differ in function and in composition: - primary walls surround growing and dividing plant cells. - secondary wall is much thicker and stronger than primary wall and accounts for most of the carbohydrate in biomass. Middle lamella is a specialize region associated with the cell walls and is shared by neighboring cells. Plasmodesmata is the small passages penetrate the middle larmella as well ass the primary and secondary cell walls, providing pathways for transporting cytoplasmic molecules from one cell to another. tainano.com/.../image036.gif

8
Plant cell wall structure Primary Cell Walls The main chemical components of the primary plant cell wall include cellulose. In addition, the cell wall contains two groups of branched polysaccharides, the pectins and cross-linking glycans. Organized into a network with the cellulose microfibrils, the cross-linking glycans increase the tensile strength of the cellulose, whereas the coextensive network of pectins provides the cell wall with the ability to resist compression. The middle lamella is rich in pectins. pectin cross-linking glycan cellulose microfibrils middle lamella primary cell wall plasma membrane micro.magnet.fsu.edu/cells/plamts/cellwall.html

9
Plant cell wall structure Secondary Cell Wall The secondary cell walls contain cellulose, hemicellulose and pectin as well as the primary cell walls, albeit in different proportion. The cellulose fibrils are embedded in a network of hemicellulose and lignin to make a further strengthened form. Numerous technical challenges must be overcome to enable the efficient utilization of secondary walls for energy production. middle lamella primary cell wall cellulose hemicellulose lignin plasma membrane protein www.ccrc.uga.edu/~mao/intro/ouline.htm

10
Structure of cellulose Cellulose is a linear condensation polymer consisting of glucose subunit linked by -1, 4-glycosidic bonds. CellobioseGlucose Coupling of adjacent cellulose molecules by the hydrogen bonds and van der Waal’s forces results in a parallel alignment and a crystalline structure. Approximately 30 individual crystalline cellulose molecules are packed into larger units called microfibrils, which are in turn assembled into the familiar cellulose fibers. Microfibril 7-30 nm Lignin Hemicellulose Elementary cellulose fibril

11
The microfibril chain are oriented in parallel and form highly ordered, crystalline domains interspersed by more disordered, amorphous regions. Structure of cellulose Crystalline domain Amorphous region The native, crystalline form of cellulose has a structure designated as type I, which can be converted to other crystalline forms (II-IV) by various treatments. Cellulose I can be transformed into cellulose II by alkali treatment and into cellulose III by supercritical ammonia treatment. Cellulose IV could be prepared by heat treatment in glycerol after transformation into cellulose II or cellulose III but cannot be transformed directly from cellulose I. Native cellulose has two distinct crystallite form, I and I , which differ in their intermolecular hydrogen bonding pattern. Form I is dominant in bacterial and algal cellulose and form I is dominant in higher plants.
 by various treatments. Cellulose I can be transformed into cellulose II by alkali treatment and into cellulose III by supercritical ammonia treatment. Cellulose IV could be prepared by heat treatment in glycerol after transformation into cellulose II or cellulose III but cannot be transformed directly from cellulose I. Native cellulose has two distinct crystallite form, I and I , which differ in their intermolecular hydrogen bonding pattern. Form I is dominant in bacterial and algal cellulose and form I is dominant in higher plants..")
12
Cellulolytic Organisms In plants, cellulases hydrolyze their cell walls at various developmental stages (e.g., bean abscission, fruit ripening and abscission, and pedicel abscission). For microorganisms and animals, hydrolysis and utilization of cellulose by cellulsas can provide usable energy to them. Microorganism - Bacteria: fermentative anaerobes (Clostridium, Ruminococcus, Fibrobacter) aerobic gram-positive (Cellulomonas, Thermobifida) aerobic gliding (Cytophaga, Sporocytophaga) Ruminococcus CellulomonasCytophagaClostridium
 aerobic gram-positive (Cellulomonas, Thermobifida) aerobic gliding (Cytophaga, Sporocytophaga) Ruminococcus CellulomonasCytophagaClostridium.")
13
Microorganism - Protozoa: ciliate (Diplodinium, Eudiplodinium) Protozoa of Diplodinium and Eidinium type attached to fodder molecules in rumen liquid (Dobicki et al., 2006) Piromyces AnaeromycesChaetomiumTrichodermaPhanerochaete - Fungi: monocentric (Neocllimastix, Piromyces, Caecomyces) policentric (Orpimomyces, Anaeromyces) Ascomycetes (Bulgaria, Chaetomium, Helotium) Basidiomycetes (Coriolus, Phanerochaete, Serpula) Deuteromyces (Aspergillus, Cladosporium, Penicillium, Trichoderma)
 Protozoa of Diplodinium and Eidinium type attached to fodder molecules in rumen liquid (Dobicki et al., 2006) Piromyces AnaeromycesChaetomiumTrichodermaPhanerochaete - Fungi: monocentric (Neocllimastix, Piromyces, Caecomyces) policentric (Orpimomyces, Anaeromyces) Ascomycetes (Bulgaria, Chaetomium, Helotium) Basidiomycetes (Coriolus, Phanerochaete, Serpula) Deuteromyces (Aspergillus, Cladosporium, Penicillium, Trichoderma)")
14
Animal - Arthropods: termite (Coptotermes, Nasutitermes, Neotermes, Reticulitermes) cockroach (Cryptocercus, Panesthia) beetle (Ergates) Symbioses - higher termite-bacteria - lower termite-bacteria-protozoa - lower termite-fungus (Ohkuma, 2003) Fungus (Termitomyces) Termite nest (Odontotermes) NasutitermesCryptocercusErgates
 cockroach (Cryptocercus, Panesthia) beetle (Ergates) Symbioses - higher termite-bacteria - lower termite-bacteria-protozoa - lower termite-fungus (Ohkuma, 2003) Fungus (Termitomyces) Termite nest (Odontotermes) NasutitermesCryptocercusErgates")
15
Animal - Molluscs snail (Achatina, Helix, Levantina) bivalve (Mytilus, Xylophaga) sea slug (Dolabella) marine periwinkle (Littorina)
 bivalve (Mytilus, Xylophaga) sea slug (Dolabella) marine periwinkle (Littorina)")
16
Cellulase Enzyme Systems Since cellulose cannot get into the cells, cellulolytic enzymes are by necessity secreted into the medium or bound to the outside surface of cellulolytic organisms. General Feature Microorganisms - cellulase systems including a multiplicity of enzyme components - cellulase systems exhibiting higher collective activity than the sum of the activities of individual enzymes - most cellulases containing both catalytic and carbohydrate- binding modules - cellulases often associated with each other and with the surface of cellulolytic microorganisms Termite - endogeneous cellulase excreted from the salivary glands or the mid-gut - cellulases of termite origin belonging to glycosyl hydrolase family 9 - cellulases containing a single catalytic domain and lacking the ancillary domains such as cellulose-binding domain - cellulase systems consisting of endogeneous cellulases from termite and cellulolytic enzymes of gut protists

17
Components of cellulase systems Cellulases are distinguished from other glycoside hydrolases by their ability to hydrolyze -1, 4-glucosidic bind between glucosyl residues. Based on mode of catalytic action and on structural properties, three major types of enzymatic activities are found: (i) Endo- -1, 4-glucanase (E.C. 3.2.1.4) cleaves at internal amorphous site in the cellulose polysaccharide chain, generating oligosaccharides of various lengths and consequently new chain end (ii) Exoglucanase (cellodextrinases E.C. 3.2.1.74 or cellobiohydrolase E.C. 3.2.1.91) acts in a processive manner on the reducing or nonreducing ends of cellulose polysaccharide chain, releasing either glucose or cellulobiose (iii) -Glucosidase (E.C. 3.2.1.21) hydrolyzes soluble cellodextrins and cellobiose to glucose
 Endo- -1, 4-glucanase (E.C ) cleaves at internal amorphous site in the cellulose polysaccharide chain, generating oligosaccharides of various lengths and consequently new chain end (ii) Exoglucanase (cellodextrinases E.C or cellobiohydrolase E.C ) acts in a processive manner on the reducing or nonreducing ends of cellulose polysaccharide chain, releasing either glucose or cellulobiose (iii) -Glucosidase (E.C ) hydrolyzes soluble cellodextrins and cellobiose to glucose.")
18
Components of cellulase systems Reaction mechanisms It is generally assumed that the hydrolysis reaction catalyzed by cellulase proceeding via an acid-base mechanism involving two residues, one as a general acid catalyst and another as a nucleophile. Asp-201 Glu-555 Catalytic mechanism of C. thermocellum endoglucanase CelD as a paradigm of -glucanase acting (Beguin and Aubert, 1992))
).")
19
Microorganism cellulase system Microorganism have adapted different approaches to effectively hydrolyze cellulose. There are two different systems: Noncomplexed cellulase systems The microorganisms, such as cellulolytic filamentous fungi and actinomycete bacteria, have the ability to penetrate cellulolytic substrate and produce ”free” cellulases, with or without cellulose- binding modules. Crystalline Amorphous Endoglucanase -glucosidase Exoglucanase CellobioseGlucose Cello-oligosaccharides Nonreducimg end Reducing end (Lynd et al., 2002)
.")
20
Microorganism cellulase system Complexed cellulase systems The microrganisms lack the ability to effectively penetrate cellulosic material and perhaps exists under a condition in the present of competition from other microorganisms and with limited ATP available for cellulase synthesis. This could have led to the development of “complexed” cellulase systems (called “cellulosome”). Crystalline Amorphous Cellobiose Endoglucanase (with dockerin) Exoglucanase (with dockerin) Exoglucanase (with dockerin) Cohesin moiety Carbohydrate-binding module (CBM) Cellobiose/cellodextrin phosphorylase Glucose Cello-oligosaccharides Bacterium cell wall Scaffodin (Lynd et al., 2002)
. Crystalline Amorphous Cellobiose Endoglucanase (with dockerin) Exoglucanase (with dockerin) Exoglucanase (with dockerin) Cohesin moiety Carbohydrate-binding module (CBM) Cellobiose/cellodextrin phosphorylase Glucose Cello-oligosaccharides Bacterium cell wall Scaffodin (Lynd et al., 2002).")
21
Microorganism cellulase system Cellulose-binding domain (CBD) CBDs provide a specific means for linking enzymes or other proteins on cellulose. These domains are usually located at the NH 2 or COOH terminus of the enzymes and are often separated from the catalytic domains by glycosylated, Pro/Thr/Ser-rich linker segments. Hydrogen bond formation and van Waals interactions are the main driving forces for binding. In proteins that possess hydrolytic activity the CBD concentrates its catalytic domains on the surface of the insoluble cellulose substrate. In proteins that have no hydrolytic activity, CBD is part of a scaffolding subunit that organizes the catalytic subunits into a cohesive multienzyme complex know as a cellulosome. Based on amino acid sequences, binding specificity and structures, CBD can be divided to 16 different families among 48 different carbohydrate-binding modules.

22
Microorganism cellulase system Cellulose-binding domain (CBD) Most of the CBDs found belong to the four major families: FamilyIIIIIIIV Typical size33-4090-108130-172125-170 Shapewedgeelongated barrelsbarrels- Binding reversible binds to surface of both amorphous and crystalline cellulose, not to soluble cellulose irreversible binds to surface of both amorphous and crystalline cellulose, not to soluble cellulose irreversible binds to surface of both amorphous and crystalline cellulose, not to soluble cellulose reversible binds to single molecules of amorphous cellulose or soluble cellulose, not to crystalline cellulose Characteristics Exclusively CBDs from fungal enzymes Two sub-families (IIa and IIb) CBDs from bacterial enzymes Two sub-families (IIIa and IIIb) CBDs from bacterial enzymes Also found in cellulososme CBDs from bacterial enzymes (Shoseyov and Warren, 1997)
 Most of the CBDs found belong to the four major families: FamilyIIIIIIIV Typical size Shapewedgeelongated barrelsbarrels- Binding reversible binds to surface of both amorphous and crystalline cellulose, not to soluble cellulose irreversible binds to surface of both amorphous and crystalline cellulose, not to soluble cellulose irreversible binds to surface of both amorphous and crystalline cellulose, not to soluble cellulose reversible binds to single molecules of amorphous cellulose or soluble cellulose, not to crystalline cellulose Characteristics Exclusively CBDs from fungal enzymes Two sub-families (IIa and IIb) CBDs from bacterial enzymes Two sub-families (IIIa and IIIb) CBDs from bacterial enzymes Also found in cellulososme CBDs from bacterial enzymes (Shoseyov and Warren, 1997)")
23
Microorganism cellulase system Cellulosome The plant cell wall degrading enzymes in most anaerobic microorganisms associate into a supramolecular complex, termed the “cellulosome” with a molecular mass higher than 2 MDa. The cellulosomes are associated with the cell surface and mediate cell attachment to the insoluble substrate and degrade it to soluble products which are then absorbed. cellulosomes Cell membrane S-layer anchoring proteins cellulosomes Cell cellulose (Bayer et al., 1998)
.")
24
Microorganism cellulase system Cellulosome The principal component of the cellulosome is a scaffoldin subunit that contains cohesin modules and also frequently includes a carbohydrate-binding module. The enzymatic subunits of the cellulosome contain a complementary type of module, the dockerin domain, that is responsible for attachment to the cohesin modules of scaffoldin. bacterial cell N N C C Type I Dockerin Type II Dockerin Type I Cohesin Type II Cohesin Scaffoldin SLH Module X Module Cellulose-binding domain Catalytic domain (Demain et al., 2005)
.")
25
Microorganism cellulase system Cellulosome Recently, several different modular structures of scaffoldin have been described in various anaerobic microognisms. 123456789 XII CBD Clostridium thermocellum CipA X X CBD 12345678 C. celluloticum CipC CBD II 54321 67891011 Bacteroides cellulosolvens ScaA 123456789 X XXX CBD C. cellulovorans CbpA GH9 1234567 XII CBD Acetivibrio cellulolyticus CipV X CBD 123456 C. josui CipJ

26
Microorganism cellulase system Cellulosome The cellulosome of most anaerobic bacteria is in essence a cell- surface component which need a cell-surface protein to mediate the its binding. The most studied specie, Clostridium thermocellum, has been identified four cell-surface proteins and one scaffoldin that are relative to this binding. Gene product Description SdbAScaffoldin-dockerin binding component A Orf2pCell surface glycoprotein 2 OlpBOuter layer protein component B OlpAOuter layer protein component A SlpAS-layer glycoprotein A SdbA Orf2p OlpB OlpA Bacterial Cell (Demain et al., 2005) (Bayer et al., 1998)
 (Bayer et al., 1998).")
27
Termite cellulase system The presence of gut protists is important to termite survival on a diet of cellulose as their energy and carbon source. Termites grind and crunch their ingested material, which may enhance digestion by increasing the amount of surface that can be accessed by cellulolytic enzymes. Probably, the ingested cellulose can be partially degraded by the endoglucanase of termite origin, and the cellulose not hydrolyzed in the anterior portion of the gut then travels to the hindgut, where it can be endocytosed and fermented by the symbiotic microrganisms. Cellulolytic material Gring Crunch Mid-gut endogeneous cellulase Ingested cellulose Hindgut Cellulose decomposition Protist CO 2 H2H2 CH 4 Acetate Absorbed by termite

28
Regulation of Cellulase Production Carbon Source Regulation Cellulose and Derived Metabolites For most microorganisms, cellulase synthesis generally requires the presence of cellulose or its soluble metabolites. Cellobiose functions as an inducer is more complex because at high level it inhibits cellulase production. In case of fungi, cellulose induces cellulase synthesis in germinating conidia, but not in mycelium. Sophorose ( -1,2-glucobiose), that is formed via the transglycosylation of cellobiose by a -glucosidase, was identified as a strong inducer of cellulase formation in fungi. Easily Metabolized Substrates In most moicroorganisms, cellulase synthesis is blocked in presence of soluble substrate such as glucose.
, that is formed via the transglycosylation of cellobiose by a -glucosidase, was identified as a strong inducer of cellulase formation in fungi. Easily Metabolized Substrates In most moicroorganisms, cellulase synthesis is blocked in presence of soluble substrate such as glucose..")
29
Intracellular Molecules Transcriptional Factors ACEI and ACEII were identified the ability to bind to the promoter region of fungal cellulase gene thus can stimulate the expression of cellulase gene. Gene Cluster In the case of anaerobic bacteria, several gene clusters have been found, suggesting the existence of operons as units of gene regulation. Negative control For clostridia, three protein, GlyR1, GlyR2, and GlyR3 were identified as regulatory proteins containing two major domains, a sugar-binding domain and a DNA-binding domain. These proteins inhibit the cellulase production by binding to the promoter region of cellulase gene.

30
Intracellular Molecules Carbon Catabolite Repressor Cre1 or CreA was identified as an inhibitor for the transcription of cellulase gene of fungus. ATP and cAMP In the case of fungi, extracellular cellulase was repressed at intracellular ATP concentration at a high level and cAMP played a role in derepression of enzyme synthesis. Enzyme Inhibitor Nojirimycin and Gluconolactone The inhibition of -glucosidase activity by these components may prevent induction by celloluse.

31
Cellulases Some cellulase enzymes may play a role in the formation of the inducer for other enzymes. In T. reesei, Fowler and Brown (1992) suggested that BGL1 may be partially responsible for formation of the inducer because the deletion of bgl1 gene resulted in decreased endoglucanase activities. Later, Seiboth et al. (1997) revealed that deletion of cbh2 and eg2 genes prevented the expression of other cellulase genes. In R. flavefaciens, Doerner et al. (1992) reported that the celA and celC genes were expressed constitutively while expression of the celB and celD was induced by cellulose. In C. thermocellum, Mishra (1991) revealed that transcription of several cel genes was induced sequentially when cellobiose concentration in the medium became limiting. Transcription of celA started first, followed by celD and celF, and finally celC.
 suggested that BGL1 may be partially responsible for formation of the inducer because the deletion of bgl1 gene resulted in decreased endoglucanase activities. Later, Seiboth et al. (1997) revealed that deletion of cbh2 and eg2 genes prevented the expression of other cellulase genes. In R. flavefaciens, Doerner et al. (1992) reported that the celA and celC genes were expressed constitutively while expression of the celB and celD was induced by cellulose. In C. thermocellum, Mishra (1991) revealed that transcription of several cel genes was induced sequentially when cellobiose concentration in the medium became limiting. Transcription of celA started first, followed by celD and celF, and finally celC..")
32
Summery Cellulose first undergoes limited hydrolysis by cellulases constitutively produced in low amounts. The soluble hydrolysis products thus generate and cause induction of cellulase synthesis. Catabolite repression of cellulase genes occurs in the presence of glucose and may be regulated by cAMP.

33
APPLICATION OF CELLULASES

34
Methodology for Studying Cellulase Properties Substrate for cellulase activity assays Substrate for cellulase activity assays can be divided into 2 categories, based on their solubility in water. SubstrateDetection a Enzymes Soluble Short chain (low DP) Cellodextrins Radio-labeled cellodextrins Cellodextrin derivatives -methylumbelliferyl –oligosaccharides p-nitrophenol-oligosaccharides Long chain cellulose derivatives Carboxymethyl cellulose (CMC) Hydroxyethyl cellulose (HEC) Dyed CMC RS, HPLC, TLC TLC plus liquid scintillation Fluorophore liberation, TLC Chromophore liberation, TLC RS, viscosity Dye liberation Endo,Exo, BG Endo a RS, reducing sugars; TSS, total soluble sugars. (Zhang et al., 2006)
 Cellodextrins Radio-labeled cellodextrins Cellodextrin derivatives -methylumbelliferyl –oligosaccharides p-nitrophenol-oligosaccharides Long chain cellulose derivatives Carboxymethyl cellulose (CMC) Hydroxyethyl cellulose (HEC) Dyed CMC RS, HPLC, TLC TLC plus liquid scintillation Fluorophore liberation, TLC Chromophore liberation, TLC RS, viscosity Dye liberation Endo,Exo, BG Endo a RS, reducing sugars; TSS, total soluble sugars. (Zhang et al., 2006).")
35
SubstrateDetection a Enzymes Insoluble Crystalline cellulose Microcrystalline cellulose (Avicel) Cotton Valonia cellulose Bacterial cellulose Amorphous cellulose Phosphorous acid swollen cellulose (PASC) Alkali-swollen cellulose (RAC) Dyed cellulose Fluorescent cellulose Chromogenic and fluorephoric derivatives Trinitrophenyl-carboxymethylcellulose (TNP-CMC) Fluram-cellulose Practical cellulose-containing substrates -cellulose pertreated lignocellulosic biomass RS, TSS, HPLC RS, TSS, HPLC, TLC Dye liberation Fluorophore liberation Chromophore liberation Fluorophore liberation HPLC, RS Total, Endo, Exo Total, Endo Total Endo Total, Endo Total Substrate for cellulase activity assays a RS, reducing sugars; TSS, total soluble sugars. (Zhang et al., 2006)
.")
36
Cellulase activity assays All existing cellulase activity assays can be divided into three types: 1) the accumulation of products after hydrolysis - reducing sugars - total sugars - glucose 2) the reduction in substrate quantity - gravimetry - chemical methods 3) the change in the physical properties of substrates - swollen factor - fiber strength - structure collapse - turbidity - viscosity
 the accumulation of products after hydrolysis - reducing sugars - total sugars - glucose 2) the reduction in substrate quantity - gravimetry - chemical methods 3) the change in the physical properties of substrates - swollen factor - fiber strength - structure collapse - turbidity - viscosity")
37
Cellulase activity assays – accumulation of hydrolysis products Reducing sugars depend on the reduction of inorganic oxidants such as cupric ions (Cu 2+ ) or ferricyanide, which accepts electrons from the donating aldehyde groups of reducing cellulose chain ends. MethodSample (ml) Reagent (ml) G amount ( g/sample) Ref. DNS Micro DNS Macro Nelson-Somogyi Micro Nelson-Somogyi Macro Nelson Semi-Micro Ferricyanide-1 Ferricyanide-2 PAHBAH Micro PAHBAH Macro BCA Modified BCA 1-3 0.5 1-5 2 1-3 1 0.5 0.01 0.5 1 3 2+2 2 1+5 0.25 1.5 3 0.5 1 20-600 100-2500 1-10 10-600 5-100 1-9 0.18-1.8 0.5-5 5-50 0.2-4.5 0.4-9 Miller, 1959 Ghose, 1987 Somogyi, 1952 Nelson, 1944 Park & Johnson, 1949 Kidby & Davidson, 1973 Lever, 1972 Waffenschmidt & Janeicke, 1987 Zhang & Lynd, 2005 G: reducing sugar; DNS: dinitrosalicyclic acid; PAHBAH: 4-hydroxybenzoylhydrazine; BCA: 2,2’-bicinchroninate The common colorimetric reducing sugar assays
 Reagent (ml) G amount ( g/sample) Ref. DNS Micro DNS Macro Nelson-Somogyi Micro Nelson-Somogyi Macro Nelson Semi-Micro Ferricyanide-1 Ferricyanide-2 PAHBAH Micro PAHBAH Macro BCA Modified BCA Miller, 1959 Ghose, 1987 Somogyi, 1952 Nelson, 1944 Park & Johnson, 1949 Kidby & Davidson, 1973 Lever, 1972 Waffenschmidt & Janeicke, 1987 Zhang & Lynd, 2005 G: reducing sugar; DNS: dinitrosalicyclic acid; PAHBAH: 4-hydroxybenzoylhydrazine; BCA: 2,2’-bicinchroninate The common colorimetric reducing sugar assays.")
38
Cellulase activity assays – accumulation of hydrolysis products Total soluble sugars Phenol or anthrone in the presence of sulfuric acid can be used for the quantitative colorimetric microdetermination of sugars and their methylderivatives, oligosaccharides, and polysaccharides, to give an orange-yellow or blue-green color. MethodSample (ml) Reagent (ml) G amount ( g/sample) Ref. Phenol-H 2 SO 4 Anthrone-H 2 SO 4 1111 1+5 5-100 Dubois et al., 1956; Zhang & Lynd, 2005 Roe, 1955; Viles & Silverman, 1949 G: reducing sugar The common colorimetric total sugar assays
 Reagent (ml) G amount ( g/sample) Ref. Phenol-H 2 SO 4 Anthrone-H 2 SO Dubois et al., 1956; Zhang & Lynd, 2005 Roe, 1955; Viles & Silverman, 1949 G: reducing sugar The common colorimetric total sugar assays.")
39
Cellulase activity assays – accumulation of hydrolysis products Glucose Assay Enzymatic Glucose Assays depend on the glucose oxidase-peroxide reaction for the determination of glucose concentrations by using coupled hexokinase (HK) and glucose-6-phosphate dehydrogenase(PGHD). MethodSample (ml) Reagent (ml) G amount ( g/sample) Ref. Glucose-HK/PGHD kit 0.01 0.5 1 0.5 2-50 Sigma kit Zhang & Lynd, 2004 G: reducing sugar The colorimetric enzymatic glucose assays HPLC After post-hydrolysis conversion to glucose
 Reagent (ml) G amount ( g/sample) Ref. Glucose-HK/PGHD kit Sigma kit Zhang & Lynd, 2004 G: reducing sugar The colorimetric enzymatic glucose assays HPLC After post-hydrolysis conversion to glucose.")
40
Cellulase activity assays – loss of substrate Gravimetry uses precipitation or volatilization method based on the determination of a substance of known composition that is chemically related to the sugar. The standard deviation of this method is strongly associated with sample weight. Chemical Method includes the phenol-H 2 SO 4 and the anthrone-H 2 SO 4 method for residual cellulose, and HPLC quantitative saccharification for different carbohydrate components.

41
Cellulase activity assays – physical cellulose properties Swollen Factor measures by alkali uptake. Structure Collapse measures the reduction in tensile strength of cellulotic fiber. Turbidity measures a reduction in the absorbance of particle suspension during the hydrolysis process. Amorphous cellulose is recommended for this assay. Viscosity measures a reduction in substrate viscosity. Soluble cellulose derivatives are recommended for this assay.

42
Endoglucanase activity assays Endoglucanases cleave intramolecular -1,4-glucosidic linkages randomly, and their activities can be measured based on a reduction in substrate viscosity and/or an increase in reducing ends determined by a reducing sugar assay. Because exoglucanases also increase the number of reducing ends, it is strongly recommended that endoglucanase activities be measured by both methods. CMC, a soluble high DP (degree of polyerization) cellulose derivative, is often recommended as a good substrate for endoglucanase activities. Soluble oligosaccharides and their chromophore-substituted substrates, such as p-nitrophenyl glucosides and methylumbelliferyl- - D -glucosides, are also to measure endoglucanase activities based on the release of chromophores or the formation of shorter oligosaccharide fragments, which are measured by HPLC or TLC. Endoglucanase activities can also be easily detected on agar plates by staining residual polysaccharides (CMC, cellulose) with various dyes, such as Congo red, because these dyes are adsorbed only by long chains of polysaccharides.
 cellulose derivative, is often recommended as a good substrate for endoglucanase activities. Soluble oligosaccharides and their chromophore-substituted substrates, such as p-nitrophenyl glucosides and methylumbelliferyl- - D -glucosides, are also to measure endoglucanase activities based on the release of chromophores or the formation of shorter oligosaccharide fragments, which are measured by HPLC or TLC. Endoglucanase activities can also be easily detected on agar plates by staining residual polysaccharides (CMC, cellulose) with various dyes, such as Congo red, because these dyes are adsorbed only by long chains of polysaccharides..")
43
Exoglucanase activity assays Exoglucanases cleave the accessible ends of cellulose molecules to liberate glucose and cellobiose. During chromatographic fractionation of cellulase mitures, enzymes with little activity on soluble CMC, but showing relatively high activity on Avicel, are usually identified as exoglucanase. There is no substrates specific for exoglucanase within the cellulase mixtures. Soluble oligosaccharides and their chromophore-substituted substrates, such as p-nitrophenyl- - D -cellobioside, 4- methylumbelliferyl- - D -lactoside, 4-methylumbelliferyl- - D - aglycones and 4-methylumbelliferyl- - D -glycosides, are also to measure exoglucanase activities, which can be differentiated from endoglucanase activities by the aid of their specific inhibitor such as cellobiose. But different exoglucanases have different activities on these substrates.

44
glucosidase activity assays -glucosidase hydrolyze soluble cellobiose and other cellodextrins in the aqueous phase. These enzymes are very amenable to a wide range of simple sensitive assay methods, based on colored or fluorescent products released from p-nitrophenyl - D -1,4- glucopyranoside, -naphthyl- - D -glucopyranoside, 6-bromo-2- naphthyl- - D -glucopyranoside, and 4-methylumbelliferyl- - D - glucopyranoside. Cellobiose, which is not hydrolyzed by endoglucanases and exoglucanases, is also used as a substrate for -glucosidase activity assays by determining the increase of reducing sugar.

45
Total cellulase activity assays The total cellulase activity assays are always measured using insoluble substrates, including pure cellulosic substrates such as Whatman No. 1 filter paper, cotton fiber, microcrystalline cellulose, bacterial cellulose, algal cellulose; and cellulose-containing substrates such as dyed cellulose, -cellulose, and pretreated lignocellulose. The most common total cellulase activity assay is filter paper assay (FPA) which requires a fixed amount (2mg) of glucose released from a 50-mg sample of Whatman No.1 filter paper. -cellulose and pretreated lignocellulose are often used to evaluate the digestibility of a reconstituted cellulase mixture for a prolonged reaction. Dyed celluloses and fluorescent-dyed celluloses are widely used for determining sugar inhibition for total cellulase.
 which requires a fixed amount (2mg) of glucose released from a 50-mg sample of Whatman No.1 filter paper. -cellulose and pretreated lignocellulose are often used to evaluate the digestibility of a reconstituted cellulase mixture for a prolonged reaction. Dyed celluloses and fluorescent-dyed celluloses are widely used for determining sugar inhibition for total cellulase..")
46
Recombinant Cellulolytic Strategy Fundamentals of microbial cellulose utilization Underlying fundamental issue: Understand cellulose hydrolysis at a microbial rather than an enzymatic level Engineered strains able to utilize cellulose and produce a desired product at high yield Microbes with good substrate utilization properties: Cellulase production, utilization of hydrolysis products (e.g. thermophiles) Microbes with good product-producing properties: High product yields, titers ect. (e.g. yeasts) Native strategy: Metabolic engineering to improve product yields, titer ect. Recombinant strategy: Heterologous cellulase expression Strain characterization and improvemetn Applied objective: CBP-compatible strains of use for industrial processes (Lynd et al., 2005)
 Microbes with good product-producing properties: High product yields, titers ect. (e.g. yeasts) Native strategy: Metabolic engineering to improve product yields, titer ect. Recombinant strategy: Heterologous cellulase expression Strain characterization and improvemetn Applied objective: CBP-compatible strains of use for industrial processes (Lynd et al., 2005).")
47
Strain-donor The most studied cellulolytic organisms and their properties are listed as follow: SpecieFunction Acidothermus cellulolyticus Clostridium thermocellum Coptotermes formosanus Erwinia chrysanthemi Humicola grisea Nasutitermes takasagoensis Reticulitermes speratus Thermoascus aurantiacus Thermobifida fusca Trichoderma reesei thermostable cellulase cellulosome animal cellulase high activity cellulase thermostable cellulase animal cellulase thermostable cellulase cellulase system

48
Strain-host The most studied host microorganisms and their functions are listed as follow: SpecieFunction Aspergillus oryzae Clostridium cellulolyticum Clostridium thermocellum Escherichia Coli Fusarium oxysporum Klebsiella oxytoca Neurospora crassa Saccharomyces cerevisiae Thermoanaerobacterium thermosaccarolyticum Thermoanaerbacterium saccharolyticum Trichoderma reesei Zymomonas mobilis overexpression fermentation overexpression and fermentation overexpression and fermentation expression Fermentation

49
Cellulase improvement Two methods are available for improving the properties of individual cellulase components:1) rational design and 2) directed evolution. Rational design is the eariliest approach to protein engineering and requires detailed knowledge of protein structure. (Zhang et al., 2006)
.")
50
Cellulase improvement Directed evolution is independent of knowledge of enzyme structure and of interactions between enzyme and substrate, and is developing tools to correctly evaluate the performance of mutants generated by recombinant DNA techniques. (Zhang et al., 2006)
.")
51
Cellulase improvement- cellulase structure

52
Figure7 Stereo representation of environment of the catalytic acid/base residue Glu412 at pH 5.6 and 6.5. The carboxylate group of Glu412 at pH5.6 is in purple and at pH 6.5 is in green. The simulated-annealing omit map density for Glu412 is contoured at 0.75 .

53
Cellulase improvement- cellulase structure The cellulosome is a macromolecular machine, whose components interact in a synergistic manner to catalyze the efficient degradation of cellulose. The cellulsome complex is composed of numerous kinds of cellulases and related enzyme subunits, which are assembled into the complex by virtue of a unique type of scaffolding subunit (scaffoldin). Each of the cellulosomal subunits consists of a multiple set of modules, two classes of wich (dockerin domains on the enzymes and cohesin domains on scaffoldin) govern the incorporation of the enzymatic subunits into the cellulosome complex. Another scaffoldin module-the cellulose-binding domain-is responsible for binding to the substrate. Some cellulosomes appear to be tethered to the cell envelop via similarly intricate, multiple- domain anchoring proteins. The assemblage is organized into dymatic polycellulosomal organelles, which adorn the cell surface. The cellulosome dictates both the binding of the cell to the substrate and its extracellular decomposition to soluble sugars, which are then taken up and assimilated by normal cellular processes.
. Each of the cellulosomal subunits consists of a multiple set of modules, two classes of wich (dockerin domains on the enzymes and cohesin domains on scaffoldin) govern the incorporation of the enzymatic subunits into the cellulosome complex. Another scaffoldin module-the cellulose-binding domain-is responsible for binding to the substrate. Some cellulosomes appear to be tethered to the cell envelop via similarly intricate, multiple- domain anchoring proteins. The assemblage is organized into dymatic polycellulosomal organelles, which adorn the cell surface. The cellulosome dictates both the binding of the cell to the substrate and its extracellular decomposition to soluble sugars, which are then taken up and assimilated by normal cellular processes..")
54
Cellulase improvement- cellulase structure FIG. 1. Schematic representation of cellulosome organization and attachment to the C. thermocellum cell surface. The scaffoldin protein of C. therocellum, shown in yellow, is composed primarily of nine copies of cohesin module, a Family-IIIa CBD and a type-II dockerin domain. The high-resolution crystal structures of the former tow domains have been solved and are shown in yellow in the insets. The cellulose-binding, planar aromatic strip of the CBD, and the putative dockerin-binding residues of the cohesin are highlighted in red. The schematic cellulosomal catalytic subunits are shown in shades of blue, green, and purple; the crystal structures of five of the known enzymes are shown in the large inset the top of the page. From left to right they are: endoglucanase A from C. cellulolyticum (PDB code 1EDG), cellobiohydrolase I from Trichroderma reesei (1CEL), endoglucanase E2 from Thermomonospora fusca (1TML), endoglucanase V from Humicola insolens (3ENG), and endoglucanase CelA from C. thermocellum (1CEM) These structures were chosen as representatives of the five classes of protein folds seen to date for the cellulosomal catalytic subunits, as designated at the top of each structure. The enxymes are shown bound to the scaffoldin protein via theirattached and highly conserved, type-I dockerin domains. A proposed structural model for the calcium-dependent dockerin domain, based on the EF-hand structure of troponin C (PDB code 5TNC), is shown modeled in green in the inset. The calcium ligands are shown as white spheres, and predicted positions of cohesin-recognition residues are color- coded red. The entire cellulosome, comprising the scaffoldin protein and the catalytic subunits, is bound to the cell surface (left) in either single or mutiple copies by interaction of its resident type-II dockerin with type-II cohesin domains of cell-surface anchoring proteins-SdbA, Orf2, and OlpB, shown in orange. Each of these components contains an SLH module, which anchors the parent protein and the attached scaffoldin, together with its complement of enzymes, to the cell surface. Another SLH-containing protein, OlpA, bears a type-I cohesin, which apparently serves to anchor a single cellulase to the cell exterior.
, cellobiohydrolase I from Trichroderma reesei (1CEL), endoglucanase E2 from Thermomonospora fusca (1TML), endoglucanase V from Humicola insolens (3ENG), and endoglucanase CelA from C. thermocellum (1CEM) These structures were chosen as representatives of the five classes of protein folds seen to date for the cellulosomal catalytic subunits, as designated at the top of each structure. The enxymes are shown bound to the scaffoldin protein via theirattached and highly conserved, type-I dockerin domains. A proposed structural model for the calcium-dependent dockerin domain, based on the EF-hand structure of troponin C (PDB code 5TNC), is shown modeled in green in the inset. The calcium ligands are shown as white spheres, and predicted positions of cohesin-recognition residues are color- coded red. The entire cellulosome, comprising the scaffoldin protein and the catalytic subunits, is bound to the cell surface (left) in either single or mutiple copies by interaction of its resident type-II dockerin with type-II cohesin domains of cell-surface anchoring proteins-SdbA, Orf2, and OlpB, shown in orange. Each of these components contains an SLH module, which anchors the parent protein and the attached scaffoldin, together with its complement of enzymes, to the cell surface. Another SLH-containing protein, OlpA, bears a type-I cohesin, which apparently serves to anchor a single cellulase to the cell exterior..")
55
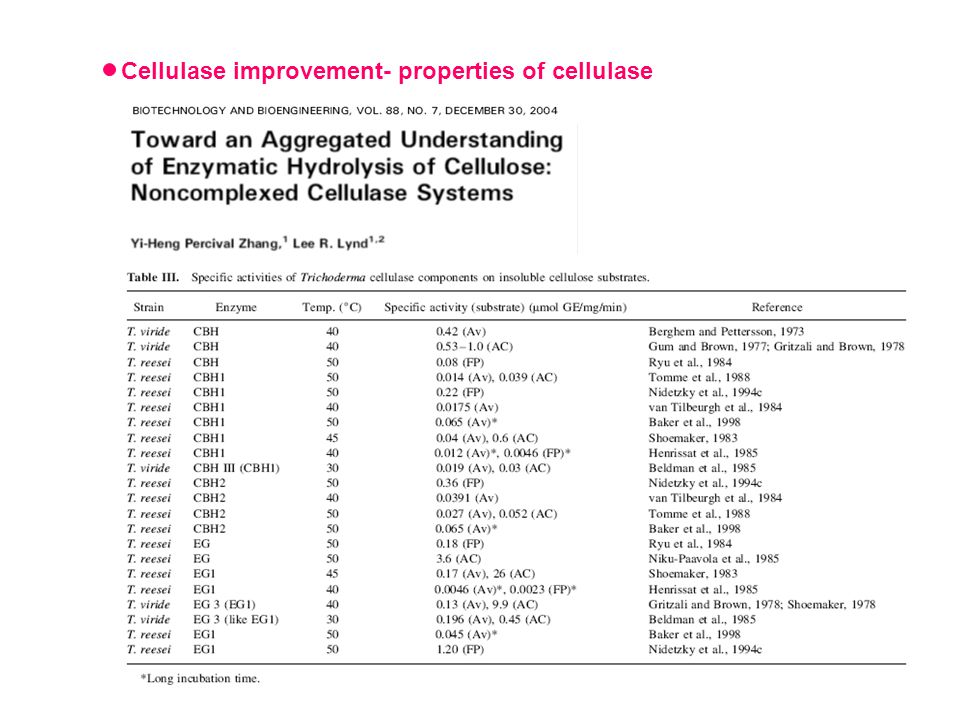
Cellulase improvement- properties of cellulase

57
G2, cellobiose; G3, cellotriose; G4, cellotetraose; G5, cellopentaose; G6, cellohexaose

58
Cellulase improvement- properties of cellulase

60
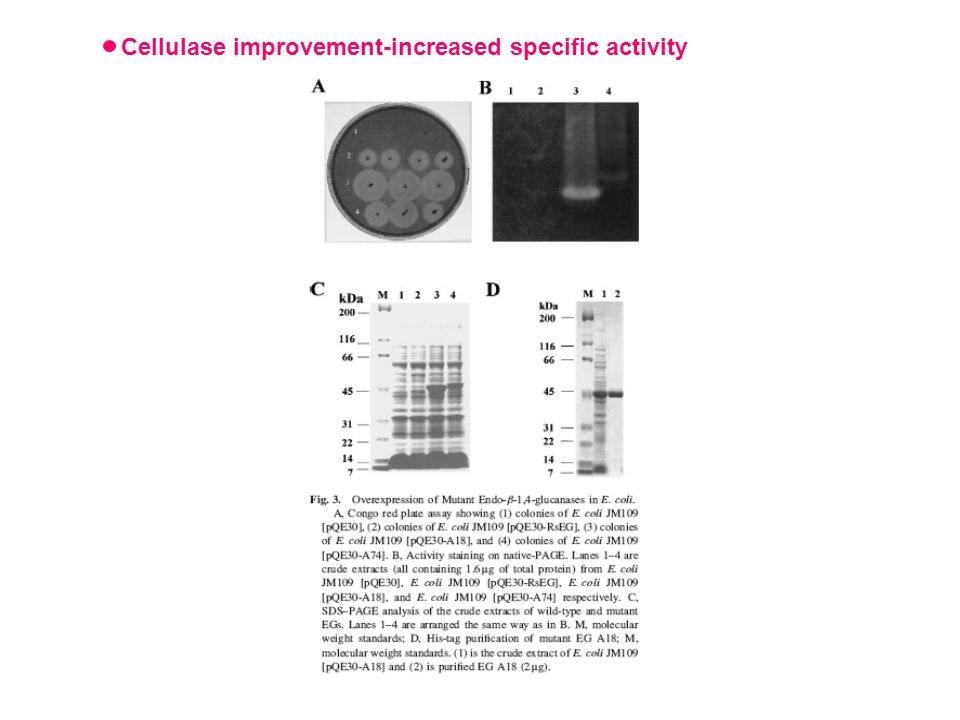
Cellulase improvement-increased specific activity

61
The level of saccharification of cellulose by T. reesei in the presence of recombinant H. grisea BGL4 was 1.4-2.2 times higher than in its absence.

62
Cellulase improvement-increased specific activity

64
Cellulase improvement-improved cellulase binding In recent work (Fierobe, H.-P., Bayer, E. A., Tardif, C., Czjzek, M., Mechaly, A., Belaïch, A., Lamed, R., Shoham, Y., and Belaich, J.-P. (2002) J. Biol. Chem. 277, 49621–49630), we reported the self-assembly of a comprehensive set of defined "bifunctional" chimeric cellulosomes. Each complex contained the following: (i) a chimeric scaffoldin possessing a cellulose-binding module and two cohesins of divergent specificity and (ii) two cellulases, each bearing a dockerin complementary to one of the divergent cohesins. This approach allowed the controlled integration of desired enzymes into a multiprotein complex of predetermined stoichiometry and topology. The observed enhanced synergy on recalcitrant substrates by the bifunctional designer cellulosomes was ascribed to two major factors: substrate targeting and proximity of the two catalytic components. In the present work, the capacity of the previously described chimeric cellulosomes was amplified by developing a third divergent cohesin- dockerin device. The resultant trifunctional designer cellulosomes were assayed on homogeneous and complex substrates (microcrystalline cellulose and straw, respectively) and found to be considerably more active than the corresponding free enzyme or bifunctional systems. The results indicate that the synergy between two prominent cellulosomal enzymes (from the family-48 and -9 glycoside hydrolases) plays a crucial role during the degradation of cellulose by cellulosomes and that one dominant family-48 processive endoglucanase per complex is sufficient to achieve optimal levels of synergistic activity. Furthermore cooperation within a cellulosome chimera between cellulases and a hemicellulase from different microorganisms was achieved, leading to a trifunctional complex with enhanced activity on a complex substrate.
 J. Biol. Chem. 277, 49621–49630), we reported the self-assembly of a comprehensive set of defined bifunctional chimeric cellulosomes. Each complex contained the following: (i) a chimeric scaffoldin possessing a cellulose-binding module and two cohesins of divergent specificity and (ii) two cellulases, each bearing a dockerin complementary to one of the divergent cohesins. This approach allowed the controlled integration of desired enzymes into a multiprotein complex of predetermined stoichiometry and topology. The observed enhanced synergy on recalcitrant substrates by the bifunctional designer cellulosomes was ascribed to two major factors: substrate targeting and proximity of the two catalytic components. In the present work, the capacity of the previously described chimeric cellulosomes was amplified by developing a third divergent cohesin- dockerin device. The resultant trifunctional designer cellulosomes were assayed on homogeneous and complex substrates (microcrystalline cellulose and straw, respectively) and found to be considerably more active than the corresponding free enzyme or bifunctional systems. The results indicate that the synergy between two prominent cellulosomal enzymes (from the family-48 and -9 glycoside hydrolases) plays a crucial role during the degradation of cellulose by cellulosomes and that one dominant family-48 processive endoglucanase per complex is sufficient to achieve optimal levels of synergistic activity. Furthermore cooperation within a cellulosome chimera between cellulases and a hemicellulase from different microorganisms was achieved, leading to a trifunctional complex with enhanced activity on a complex substrate..")
65
Cellulase improvement-improved cellulase binding

67
Cellulase improvement-increased thermostability

69
Fig. 3. Thermal inactivation curve of the wild-type Clostridium thermocellum cellulase C ( ■, t 1/2 =3.28 min) and the disulfide+mutant (X, t 1/2 =9.85 min). t 1/2 is the time while the enzyme loses 50% of its activity during incubation at 70 ℃. Incubations were performed in 20 mM tris buffer, pH 7.2 at 70 ℃. The measurements were carried out at 60 ℃ in pH 6.0 succinate buffer, containing 0.2 mM pNPC. The final protein concentration was 0.008 mg/ml. Fig. 4. Thermal inactivation curve of the wild-type Clostridium thermocellum cellulase C ( ■, T 1/2 =68.0 ℃ ) and the disulfide+mutant (X, T 1/2 =69.9 ℃ ). T 1/2 is the time while the enzyme loses 50% of its activity during 10-min incubation Incubations were performed in 20 mM tris buffer, pH 7.2 for 10 min at increasing temperatures. The measurements were carried out at 60 ℃ in 100 mM succinate buffer at pH 6.0, containing 0.2 mM pNPC. The final protein concentration was 0.008 mg/ml.
 and the disulfide+mutant (X, t 1/2 =9.85 min). t 1/2 is the time while the enzyme loses 50% of its activity during incubation at 70 ℃. Incubations were performed in 20 mM tris buffer, pH 7.2 at 70 ℃. The measurements were carried out at 60 ℃ in pH 6.0 succinate buffer, containing 0.2 mM pNPC. The final protein concentration was mg/ml. Fig. 4. Thermal inactivation curve of the wild-type Clostridium thermocellum cellulase C ( ■, T 1/2 =68.0 ℃ ) and the disulfide+mutant (X, T 1/2 =69.9 ℃ ). T 1/2 is the time while the enzyme loses 50% of its activity during 10-min incubation Incubations were performed in 20 mM tris buffer, pH 7.2 for 10 min at increasing temperatures. The measurements were carried out at 60 ℃ in 100 mM succinate buffer at pH 6.0, containing 0.2 mM pNPC. The final protein concentration was mg/ml..")
70
CLOSING COMMENT After 20 years of research and development, application of cellulase system during production of bioethanol is becoming a reality. But reducing the cost of cellulase enzyme production is still a problem in the establishment of commercial process. Genetic techniques are one solution to create new cellulase production systems with possible improvement of enzyme yield and activity. It will be interesting to see whether future approaches will be: to identify new cellulases with more powerful ability to hydrolyze cellulose materials from cellulolytic organisms to improve present cellulases properties such as thermostablity, specific activity, cellulose-binding ability or synergistic action to establish a single microorganism strain or microbial system which is able to utilize both cellulose and fermentable compound

71
CLOSING COMMENT (Bayer et al., 2007)
")
Similar presentations



















 High DP Cello-oligosaccharides (HD-COS) Not picked in chromatograph Introduction Various forms.>")


