Download presentation
Presentation is loading. Please wait.

1
Computational Biology, Part 8 Protein Coding Regions Robert F. Murphy Copyright 1996-2009. All rights reserved.

2
Sequence Analysis Tasks Finding protein coding regions

3
Goal Given a DNA or RNA sequence, find those regions that code for protein(s) Given a DNA or RNA sequence, find those regions that code for protein(s) Direct approach: Look for stretches that can be interpreted as protein using the genetic code Statistical approaches: Use other knowledge about likely coding regions
 Given a DNA or RNA sequence, find those regions that code for protein(s) Direct approach: Look for stretches that can be interpreted as protein using the genetic code Statistical approaches: Use other knowledge about likely coding regions")
4
Direct Approach

5
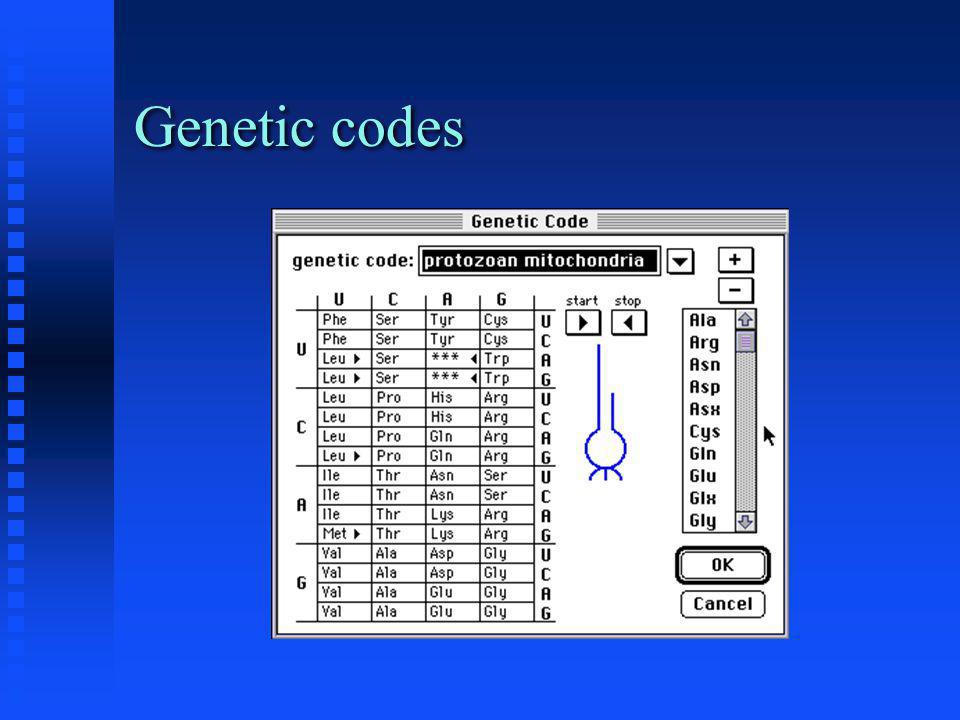
Genetic codes The set of tRNAs that an organism possesses defines its genetic code(s) The set of tRNAs that an organism possesses defines its genetic code(s) The universal genetic code is common to all organisms The universal genetic code is common to all organisms Prokaryotes, mitochondria and chloroplasts often use slightly different genetic codes Prokaryotes, mitochondria and chloroplasts often use slightly different genetic codes More than one tRNA may be present for a given codon, allowing more than one possible translation product More than one tRNA may be present for a given codon, allowing more than one possible translation product
 The set of tRNAs that an organism possesses defines its genetic code(s) The universal genetic code is common to all organisms The universal genetic code is common to all organisms Prokaryotes, mitochondria and chloroplasts often use slightly different genetic codes Prokaryotes, mitochondria and chloroplasts often use slightly different genetic codes More than one tRNA may be present for a given codon, allowing more than one possible translation product More than one tRNA may be present for a given codon, allowing more than one possible translation product")
6
Genetic codes Differences in genetic codes occur in start and stop codons only Differences in genetic codes occur in start and stop codons only Alternate initiation codons: codons that encode amino acids but can also be used to start translation (GUG, UUG, AUA, UUA, CUG) Alternate initiation codons: codons that encode amino acids but can also be used to start translation (GUG, UUG, AUA, UUA, CUG) Suppressor tRNA codons: codons that normally stop translation but are translated as amino acids (UAG, UGA, UAA) Suppressor tRNA codons: codons that normally stop translation but are translated as amino acids (UAG, UGA, UAA)
 Alternate initiation codons: codons that encode amino acids but can also be used to start translation (GUG, UUG, AUA, UUA, CUG) Suppressor tRNA codons: codons that normally stop translation but are translated as amino acids (UAG, UGA, UAA) Suppressor tRNA codons: codons that normally stop translation but are translated as amino acids (UAG, UGA, UAA)")
7
Genetic codes

9
Note additional start codons: UUA, UUG, CUG Note additional start codons: UUA, UUG, CUG Note conversion of stop codon UGA (opal) to Trp Note conversion of stop codon UGA (opal) to Trp
 to Trp Note conversion of stop codon UGA (opal) to Trp")
10
Reading Frames Since nucleotide sequences are “read” three bases at a time, there are three possible “frames” in which a given nucleotide sequence can be “read” (in the forward direction) Since nucleotide sequences are “read” three bases at a time, there are three possible “frames” in which a given nucleotide sequence can be “read” (in the forward direction) Taking the complement of the sequence and reading in the reverse direction gives three more reading frames Taking the complement of the sequence and reading in the reverse direction gives three more reading frames
 Since nucleotide sequences are read three bases at a time, there are three possible frames in which a given nucleotide sequence can be read (in the forward direction) Taking the complement of the sequence and reading in the reverse direction gives three more reading frames Taking the complement of the sequence and reading in the reverse direction gives three more reading frames")
11
Reading frames TTC TCA TGT TTG ACA GCT TTC TCA TGT TTG ACA GCT RF1 Phe Ser Cys Leu Thr Ala> RF2 Ser His Val *** Gln Leu> RF3 Leu Met Phe Asp Ser> AAG AGT ACA AAC TGT CGA AAG AGT ACA AAC TGT CGA RF4 <Glu *** Thr Gln Cys Ser RF5 <Glu His Lys Val Ala RF6 <Arg Met Asn Ser Leu

12
Open Reading Frames (ORF) Concept: Region of DNA or RNA sequence that could be translated into a peptide sequence (open refers to absence of stop codons) Concept: Region of DNA or RNA sequence that could be translated into a peptide sequence (open refers to absence of stop codons) Prerequisite: A specific genetic code Prerequisite: A specific genetic code Definition: Definition: (start codon) (amino acid coding codon) n (stop codon) Note: Not all ORFs are actually used Note: Not all ORFs are actually used
 Concept: Region of DNA or RNA sequence that could be translated into a peptide sequence (open refers to absence of stop codons) Concept: Region of DNA or RNA sequence that could be translated into a peptide sequence (open refers to absence of stop codons) Prerequisite: A specific genetic code Prerequisite: A specific genetic code Definition: Definition: (start codon) (amino acid coding codon) n (stop codon) Note: Not all ORFs are actually used Note: Not all ORFs are actually used")
14
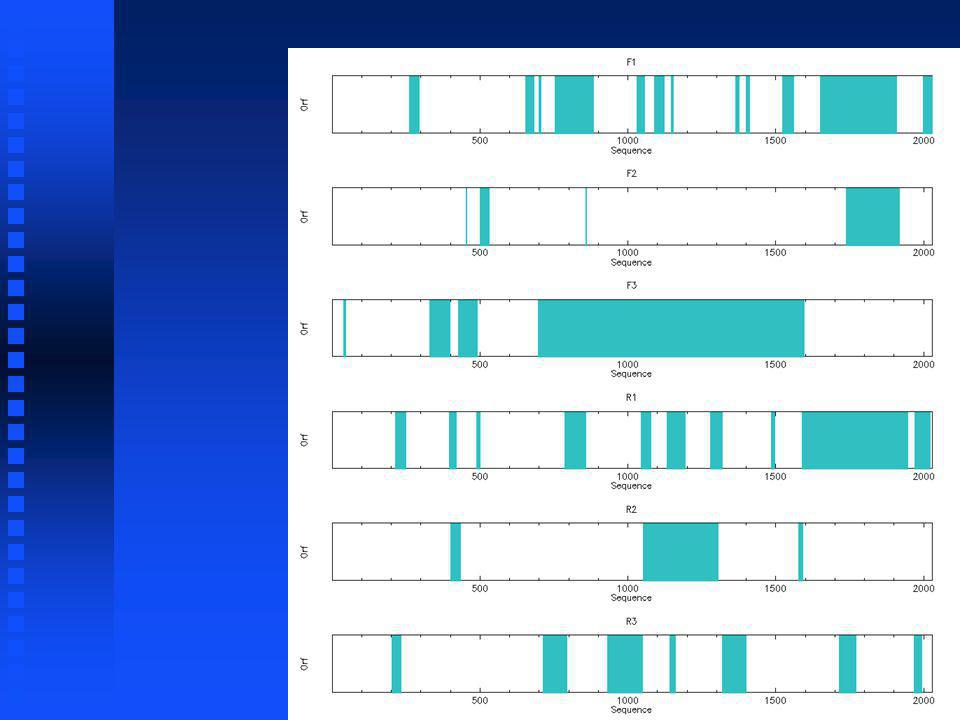
Open Reading Frames Click boxes for List ORFS and ORF map Click boxes for List ORFS and ORF map

15
Check reading frame: mod(696,3)=0 -> RF3 Check reading frame: mod(696,3)=0 -> RF3
=0 -> RF3 Check reading frame: mod(696,3)=0 -> RF3")
16
EMBOSS plotorf

19
Splicing ORFs For eukaryotes, which have interrupted genes, ORFs in different reading frames may be spliced together to generate final product For eukaryotes, which have interrupted genes, ORFs in different reading frames may be spliced together to generate final product ORFs from forward and reverse directions cannot be combined ORFs from forward and reverse directions cannot be combined

20
Block Diagram for Search for ORFs Search Engine Sequence to be searched Genetic code List of ORF positions Both strands? Ends start/stop?

21
Statistical Approaches

22
Calculation Windows Many sequence analyses require calculating some statistic over a long sequence looking for regions where the statistic is unusually high or low Many sequence analyses require calculating some statistic over a long sequence looking for regions where the statistic is unusually high or low To do this, we define a window size to be the width of the region over which each calculation is to be done To do this, we define a window size to be the width of the region over which each calculation is to be done Example: %AT Example: %AT

23
Base Composition Bias For a protein with a roughly “normal” amino acid composition, the first 2 positions of all codons will be about 50% GC For a protein with a roughly “normal” amino acid composition, the first 2 positions of all codons will be about 50% GC If an organism has a high GC content overall, the third position of all codons must be mostly GC If an organism has a high GC content overall, the third position of all codons must be mostly GC Useful for prokaryotes Useful for prokaryotes Not useful for eukaryotes due to large amount of noncoding DNA Not useful for eukaryotes due to large amount of noncoding DNA

24
Fickett’s statistic Also called TestCode analysis Also called TestCode analysis Looks for asymmetry of base composition Looks for asymmetry of base composition Strong statistical basis for calculations Strong statistical basis for calculations Method: Method: For each window on the sequence, calculate the base composition of nucleotides 1, 4, 7..., then of 2, 5, 8..., and then of 3, 6, 9... Calculate statistic from resulting three numbers

25
Codon Bias (Codon Preference) Principle Principle Different levels of expression of different tRNAs for a given amino acid lead to pressure on coding regions to “conform” to the preferred codon usage Non-coding regions, on the other hand, feel no selective pressure and can drift
 Principle Principle Different levels of expression of different tRNAs for a given amino acid lead to pressure on coding regions to conform to the preferred codon usage Non-coding regions, on the other hand, feel no selective pressure and can drift")
26
Codon Bias (Codon Preference) Starting point: Table of observed codon frequencies in known genes from a given organism Starting point: Table of observed codon frequencies in known genes from a given organism best to use highly expressed genes Method Method Calculate “coding potential” within a moving window for all three reading frames Look for ORFs with high scores
 Starting point: Table of observed codon frequencies in known genes from a given organism Starting point: Table of observed codon frequencies in known genes from a given organism best to use highly expressed genes Method Method Calculate coding potential within a moving window for all three reading frames Look for ORFs with high scores")
27
Codon Bias (Codon Preference) Works best for prokaryotes or unicellular eukaryotes because for multicellular eukaryotes, different pools of tRNA may be expressed at different stages of development in different tissues Works best for prokaryotes or unicellular eukaryotes because for multicellular eukaryotes, different pools of tRNA may be expressed at different stages of development in different tissues may have to group genes into sets Codon bias can also be used to estimate protein expression level Codon bias can also be used to estimate protein expression level
 Works best for prokaryotes or unicellular eukaryotes because for multicellular eukaryotes, different pools of tRNA may be expressed at different stages of development in different tissues Works best for prokaryotes or unicellular eukaryotes because for multicellular eukaryotes, different pools of tRNA may be expressed at different stages of development in different tissues may have to group genes into sets Codon bias can also be used to estimate protein expression level Codon bias can also be used to estimate protein expression level")
28
Portion of D. melanogaster codon frequency table

29
Comparison of Glycine codon frequencies

30
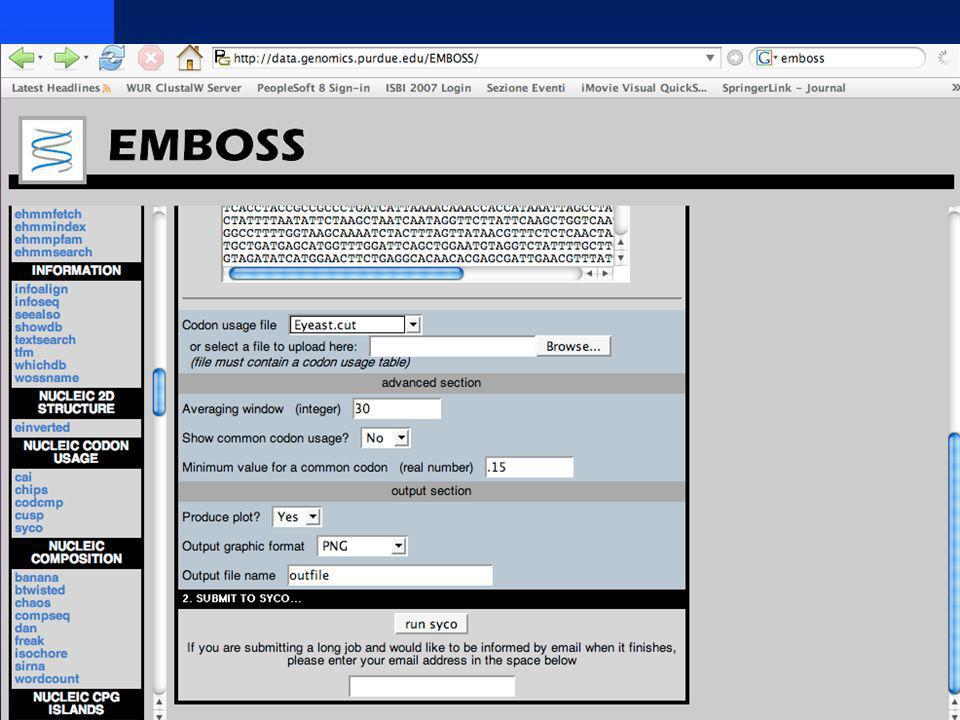
Illustration of Codon Bias Plots Use Entrez via MacVector to get sequence of lexA Use Entrez via MacVector to get sequence of lexA under “Database” select “Internet Entrez Search” Select gene=lexA AND organism=Escherichia Pick one (e.g., region from 89.2 to 92.8) Under “Analyze” select “Codon Preference Plots” Under “Analyze” select “Codon Preference Plots” Choose Escherichia coli codon bias file Choose gene region corresponding to lacZ Click on Staden codon bias and Gribskov codon bias
 Under Analyze select Codon Preference Plots Under Analyze select Codon Preference Plots Choose Escherichia coli codon bias file Choose gene region corresponding to lacZ Click on Staden codon bias and Gribskov codon bias")
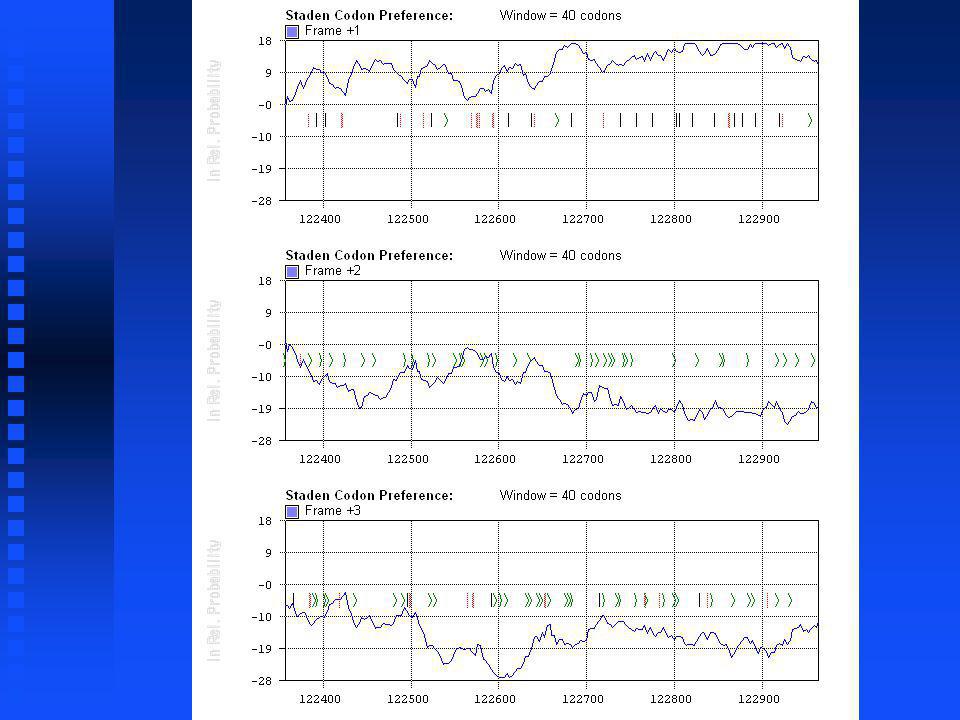
34
Codon Preference Algorithms The Staden method (from Staden & McLachlan, 1982) uses a codon usage table directly in identifying coding regions. The codon usage table is normalized so that the sum of all 64 codons is 1. The usages for each codon in each reading frame in each window are multiplied together and normalized by the sum of the probabilities in all three positions to generate a relative coding probability. The Staden method (from Staden & McLachlan, 1982) uses a codon usage table directly in identifying coding regions. The codon usage table is normalized so that the sum of all 64 codons is 1. The usages for each codon in each reading frame in each window are multiplied together and normalized by the sum of the probabilities in all three positions to generate a relative coding probability.
 uses a codon usage table directly in identifying coding regions. The codon usage table is normalized so that the sum of all 64 codons is 1. The usages for each codon in each reading frame in each window are multiplied together and normalized by the sum of the probabilities in all three positions to generate a relative coding probability..")
35
Codon Preference Algorithms The Gribskov method uses a codon usage table normalized so that the sum of the alternatives for each amino acid add to 1. The values for each codon for each reading frame in each window are multiplied together and normalized by the random probability expected for that codon given the mononucleotide frequencies of the target sequence. It is the most commonly used method. The Gribskov method uses a codon usage table normalized so that the sum of the alternatives for each amino acid add to 1. The values for each codon for each reading frame in each window are multiplied together and normalized by the random probability expected for that codon given the mononucleotide frequencies of the target sequence. It is the most commonly used method.

38
Plot from Plot from syco

39
Summary Translation of nucleic acid sequences into hypothetical protein sequences requires a genetic code Translation of nucleic acid sequences into hypothetical protein sequences requires a genetic code Translation can occur in three forward and three reverse reading frames Translation can occur in three forward and three reverse reading frames Open reading frames are regions that can be translated without encountering a stop codon Open reading frames are regions that can be translated without encountering a stop codon

40
Summary The likelihood that a particular open reading frames is in fact a coding region (actually made into protein) can be estimated using third-codon base composition or codon preference tables The likelihood that a particular open reading frames is in fact a coding region (actually made into protein) can be estimated using third-codon base composition or codon preference tables This can be used to scan long sequences for possible coding regions This can be used to scan long sequences for possible coding regions
 can be estimated using third-codon base composition or codon preference tables The likelihood that a particular open reading frames is in fact a coding region (actually made into protein) can be estimated using third-codon base composition or codon preference tables This can be used to scan long sequences for possible coding regions This can be used to scan long sequences for possible coding regions")
Similar presentations





 Translation Ribosome.>")




 (Translation) DNA (genetic information stored in genes) RNA (working copies of genes) Proteins (functional.>")








 Third mRNA base (3 end of codon)>")



. Overview of translation i). Requirements for protein synthesis ii). messenger RNA iii). Ribosomes and polysomes.>")
