Download presentation
Presentation is loading. Please wait.

1
Improvement of Industrial Strains

2
Targets for industrial strain improvement
Increase product concentration: (1) raise the gene dose; (2) break down the gene regulation (e.g. catabolite derepression, metabolite resistance); or (3) alter permeability to improve product export Process improvement: (1) decrease fermentation time; (2) be able to metabolize inexpensive substrates; (3) do not produce undesirable by products e.g. pigments or substance chemically related to the main product; (4) reduce oxygen needs; (5) decrease foaming; (6) tolerant to high concentrations of carbon or nitrogen sources; (7) resistant to phage … New product: Changes in the genotype of microorganisms can lead to the biosynthesis of new metabolites.
 raise the gene dose; (2) break down the gene regulation (e.g. catabolite derepression, metabolite resistance); or (3) alter permeability to improve product export. Process improvement: (1) decrease fermentation time; (2) be able to metabolize inexpensive substrates; (3) do not produce undesirable by products e.g. pigments or substance chemically related to the main product; (4) reduce oxygen needs; (5) decrease foaming; (6) tolerant to high concentrations of carbon or nitrogen sources; (7) resistant to phage … New product: Changes in the genotype of microorganisms can lead to the biosynthesis of new metabolites.")
3
Successful strain improvement
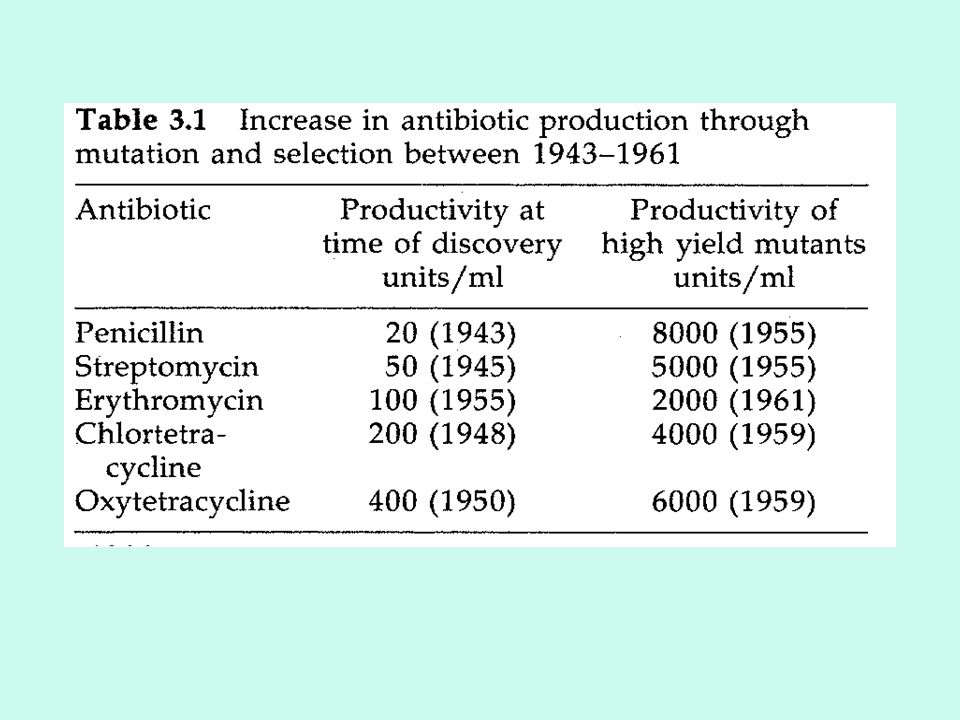
In a balanced strain development program each method should complement the other. In the past, the successes are basically due to the extensive application of mutation and selection. Current data on the production levels of industrial high-performance mutants are rarely published. Today the penicillin yield is around 85,000 units /ml (approx. 50 g/l). Because of the low yield per weight of substrate used (weight of penicillin produced per weight of glucose used is around 0.12), continued increases can be expected in the future.
. Because of the low yield per weight of substrate used (weight of penicillin produced per weight of glucose used is around 0.12), continued increases can be expected in the future.")
5
Methods used in strain improvement - 1
In vivo mutagenesis: spontaneous mutations; radiation (short-wavelength uv, nm, opt. 254nm / ionizing radiation, e.g. X-rays, -rays, and -rays); chemical agents (mutagens which affect nonreplicating DNA, e.g. nitrous acid (HNO2), hydroxylamine (NH2OH), alkylating agents / base analogs or frameshift mutagens, which are incorporated into or enter into replicating DNA; transposon insertion suicide vector and recombination
; chemical agents (mutagens which affect nonreplicating DNA, e.g. nitrous acid (HNO2), hydroxylamine (NH2OH), alkylating agents / base analogs or frameshift mutagens, which are incorporated into or enter into replicating DNA; transposon insertion. suicide vector and recombination.")
6
Methods used in strain improvement - 2
In vitro mutagenesis: random mutagenesis; directed mutagenesis (deletions / insertions / point mutations

7
Methods used in strain improvement - 3
Recombination : recombination in fungi: sexual and parasexual cycles; recombination in bacteria: transformation, transduction and conjugation; in vivo rearrangements by transposable genetic elements; protoplast fusion; in vitro recombinant DNA techniques

8
Spontaneous mutations
Mutation rate depends on growth conditions: to 10-5 per generation and per gene; usually 10-7 to 10-6. All mutant types are found, although deletions are relatively frequent. Causes include integration and excision of transposons, errors in the functioning of enzymes such as DNA polymerases, recombination enzymes, and DNA repair enzymes. Not cost-effective to isolate mutants for industrial strain development. (low frequency of mutation) The mutation frequency (proportion of mutants in the population) can be significantly increased by using mutagenic agents (mutagens): may increase to for the isolation of improved secondary metabolite producers or even up to for the isolation of auxotrophic mutants.
 The mutation frequency (proportion of mutants in the population) can be significantly increased by using mutagenic agents (mutagens): may increase to for the isolation of improved secondary metabolite producers or even up to for the isolation of auxotrophic mutants.")
9
Mutagenesis through radiation by short-wavelength ultraviolet (UV)
Effective wavelengths: nm (opt. at 254 nm) Important products: dimers between adjacent pyri-midines (T-T, T-C, and C-C) or between pyrimidines of complementary strands (results in crosslinks) Mainly induces transitions of GC→AT, transversions, frameshift and deletions are also found. To prevent error-free mechanisms of photoreactiva-tion and excision repair: carrying out all manipulation under long-wavelength visible light ( > 600 nm) and/or the use of caffeine or similar inhibitors of repair. The SOS repair system is primarily responsible for the production of mutations.
 Important products: dimers between adjacent pyri-midines (T-T, T-C, and C-C) or between pyrimidines of complementary strands (results in crosslinks) Mainly induces transitions of GC→AT, transversions, frameshift and deletions are also found. To prevent error-free mechanisms of photoreactiva-tion and excision repair: carrying out all manipulation under long-wavelength visible light ( > 600 nm) and/or the use of caffeine or similar inhibitors of repair. The SOS repair system is primarily responsible for the production of mutations.")
10
Alkylating agents Most potent mutagen for practical application, except UV Compounds frequently used: ethyl methanesulfonate (EMS), methyl methanesulfonate (MMS), diethylsulfate (DES), diepoxybutane (DEB), N-methyl-N'-nitro-N-nitrosoguanidine (NTG, the most effective chemical mutagens, but its use in a mutation program is difficult because of its carcinogenic effects), N-methyl-N-nitroso-urea and mustard gas Formation of alkylated bases in DNA, along with phosphotriester, purine-free sites and single-strand breaks / 7-alkylguanine is the most common product but it does not result in mutations / O6-alkyl-guanine and O4-alkylthymine are the most important premutational lesions, and as a result of pairing errors, mainly AT→GC transitions are elicited (direct mutagenesis). Transitions, transversions, deletions and frameshift mutations A second process results in mutation is the induction of error-prone SOS repair when relatively high doses of mutagen are used.
, methyl methanesulfonate (MMS), diethylsulfate (DES), diepoxybutane (DEB), N-methyl-N -nitro-N-nitrosoguanidine (NTG, the most effective chemical mutagens, but its use in a mutation program is difficult because of its carcinogenic effects), N-methyl-N-nitroso-urea and mustard gas. Formation of alkylated bases in DNA, along with phosphotriester, purine-free sites and single-strand breaks / 7-alkylguanine is the most common product but it does not result in mutations / O6-alkyl-guanine and O4-alkylthymine are the most important premutational lesions, and as a result of pairing errors, mainly AT→GC transitions are elicited (direct mutagenesis). Transitions, transversions, deletions and frameshift mutations. A second process results in mutation is the induction of error-prone SOS repair when relatively high doses of mutagen are used.")
11
NTG A large proportion of mutants under optimal conditions with a low killing rate. 8-10% of the survival Streptomy-ces coelicolor are auxotrophs, ~ 50% of the survival E. coli population consists of mutants 90% mutations are GC→AT transitions; also small extent deletions and frameshift after the deletion of GC pairs Easily decomposed in vivo; formed nitrous acid in acidic solutions (not effective as mutagen in pH 6-9 where NTG is active; diazomethane (a strongly methylating agent) is formed under alkaline conditions Alkylation of nonreplicating DNA and the main point of action of at the replication point of DNA, through a change in DNA polymerase III during DNA replication (there is incorrect duplication in a short segment of the DNA until the defective polymerase is replaced by an intact molecule). This explains that NTG mutations frequently occur in gene clusters.
 is formed under alkaline conditions. Alkylation of nonreplicating DNA and the main point of action of at the replication point of DNA, through a change in DNA polymerase III during DNA replication (there is incorrect duplication in a short segment of the DNA until the defective polymerase is replaced by an intact molecule). This explains that NTG mutations frequently occur in gene clusters.")
12
IS-Elements DNA sequences of variable length ( base pairs), can incorporated in different sites of the genome and released again ( recA-independent recombination) Destroy the function of the gene at the site of their integration. IS2 bears a promotor, which, when incorporated in the appropriate orientation, results in the constitutive expression of genes located downstream. In particular IS1 causes deletions, whereas IS2 causes duplications.
, can incorporated in different sites of the genome and released again ( recA-independent recombination) Destroy the function of the gene at the site of their integration. IS2 bears a promotor, which, when incorporated in the appropriate orientation, results in the constitutive expression of genes located downstream. In particular IS1 causes deletions, whereas IS2 causes duplications.")
13
Transposons Genetic elements containing flanking IS-elements in inverse orientation, often with antibiotic-resistance genes Available for a wide variety of purposes in gene technology: Tn5 contains an aminoglycoside antibiotic-resistance gene which can be expressed in a wide variety of both procaryotes and eucaryotes. Several transposons have been integrated into plasmids (e.g. Tn1 and Tn3), others in either plasmids or chromosomes (e.g. Tn5). Transposon mutagenesis offers a wide variety of advantages: (1) can obtained a mutant phenotype with a very low reversion rate, (2) relatively easy to isolate insertion mutations, (3) the site at which the transposon has been integrated can be readily determined
, others in either plasmids or chromosomes (e.g. Tn5). Transposon mutagenesis offers a wide variety of advantages: (1) can obtained a mutant phenotype with a very low reversion rate, (2) relatively easy to isolate insertion mutations, (3) the site at which the transposon has been integrated can be readily determined.")
14
Phenotypic expression of mutations - 1
Many mutations which result in increased formation of metabolites are recessive. (a) When a recessive mutation takes place in a uninuclear, haploid cell (e.g. bacteria and actinomycete spores, asexual conidia of fungi), a heteroduplex results from it; the mutant phenotype can only be expressed after a further growth step.
 When a recessive mutation takes place in a uninuclear, haploid cell (e.g. bacteria and actinomycete spores, asexual conidia of fungi), a heteroduplex results from it; the mutant phenotype can only be expressed after a further growth step.")
15
Phenotypic expression of mutations - 2
(b) Exponentially growing bacterial cells can contain 2-8 chromosomes; pure mutant clones appear until several steps in reproduction have taken place. Special procedures for filamentous actinomycetes when they lose the sporulation ability in the course of strain development: (1) grew heterokaryotic mycelium which results from mutagenesis, fragmented (ultrasonic treatment or shaking with glass beads), filtrated (paper, cotton, or an 8 m membrane filter), plated myce1ium fragments containing only one or a few nuclear bodies. Repeating this segregation process to select homokaryotic material. Or (2) the preparation of protoplasts containing one or few nuclei.
 Exponentially growing bacterial cells can contain 2-8 chromosomes; pure mutant clones appear until several steps in reproduction have taken place. Special procedures for filamentous actinomycetes when they lose the sporulation ability in the course of strain development: (1) grew heterokaryotic mycelium which results from mutagenesis, fragmented (ultrasonic treatment or shaking with glass beads), filtrated (paper, cotton, or an 8 m membrane filter), plated myce1ium fragments containing only one or a few nuclear bodies. Repeating this segregation process to select homokaryotic material. Or (2) the preparation of protoplasts containing one or few nuclei.")
16
Phenotypic expression of mutations - 3
(c) In diploid or heterokaroytic eucaryotes, recessive mutations are allowed to undergo phenotypic expression after meiosis, haploidization, or mitotic recombination. (d) Delays in expression which are not directly the result of genetic effects are observed, such as mutations which cause changed ribosomes, or (e) Mutations resulting in the loss of surface receptors (as in the development of bacteriophage resistance). In both cases, the wild type structures must be diluted out during growth before the mutation is recognizable phenotypically.
 In diploid or heterokaroytic eucaryotes, recessive mutations are allowed to undergo phenotypic expression after meiosis, haploidization, or mitotic recombination. (d) Delays in expression which are not directly the result of genetic effects are observed, such as mutations which cause changed ribosomes, or. (e) Mutations resulting in the loss of surface receptors (as in the development of bacteriophage resistance). In both cases, the wild type structures must be diluted out during growth before the mutation is recognizable phenotypically.")
17
Phenotypic expression of mutations

18
Factors affecting the appearance of mutants
The base sequence of the gene to be mutated: Different mutagenes cause hot spots (areas with high mutation frequency ) at different sites in the genome. The repair systems of the cell: Strains with partially defective repair mechanisms may be killed, so that specific mutagenes can be ineffective. Suppressor mutation: A gene activity which has become lost through mutation can be restored at least partially through a second mutation. (1)Intragenic suppressors: Exchange of an amino acid or correction of a primary frameshift mutation through deletion or insertion. (2)Extragenic suppressors: Formation of mutant transfer RNAs or ribosomes. Treatment conditions: Factors as the pH, buffer composition, mutagen concentration, exposure time, temperature, and growth phase of the organism
 at different sites in the genome. The repair systems of the cell: Strains with partially defective repair mechanisms may be killed, so that specific mutagenes can be ineffective. Suppressor mutation: A gene activity which has become lost through mutation can be restored at least partially through a second mutation. (1)Intragenic suppressors: Exchange of an amino acid or correction of a primary frameshift mutation through deletion or insertion. (2)Extragenic suppressors: Formation of mutant transfer RNAs or ribosomes. Treatment conditions: Factors as the pH, buffer composition, mutagen concentration, exposure time, temperature, and growth phase of the organism.")
19
Optimizing mutagenesis
Dose-response (killing) curves A high death rate alone is no guarantee of the occurrence of mutations in specific genes. Easily detectable changes such as mutations for resistance or re-version to auxotrophy are frequently used to optimize conditions for mutagenesis.
 curves. A high death rate alone is no guarantee of the occurrence of mutations in specific genes. Easily detectable changes such as mutations for resistance or re-version to auxotrophy are frequently used to optimize conditions for mutagenesis.")
21
Selection of improved mutants
Random selection Directed selection Rational selection

22
Selection of improved mutants - 1
Random selection: (1) 5-10 best strains are repeatedly mutated and selected; (2) Hundreds to thousands of isolates per mutation cycle must be tested (to screen a small number of survivors, about 20-50, after many different mutagen treatments and to continue mutating strains having small yield increases as quickly as possible is more economical); (3) With nonautomatic methods the number of isolates that can be tested per unit times is usually limited to per week; (4) Two stages of screening (one fermentation sample per isolation in the first stage when the test error is smaller than the yield increase expected; 10-30% best isolates are tested in a second stage and the number of replicates are chosen statistically)
 5-10 best strains are repeatedly mutated and selected; (2) Hundreds to thousands of isolates per mutation cycle must be tested (to screen a small number of survivors, about 20-50, after many different mutagen treatments and to continue mutating strains having small yield increases as quickly as possible is more economical); (3) With nonautomatic methods the number of isolates that can be tested per unit times is usually limited to per week; (4) Two stages of screening (one fermentation sample per isolation in the first stage when the test error is smaller than the yield increase expected; 10-30% best isolates are tested in a second stage and the number of replicates are chosen statistically)")
24
Selection of improved mutants - 2
Directed selection:

25
Selection of improved mutants - 3
Rational selection: (1) Antibiotic-resistant mutants: genetic marker / increased cell permeability / protein synthesis with a higher turnover; (2) Antimetabolite-resistant mutants: analog-resistant mutants can form an excess of metabolites, in some cases through changed regulatory mechanisms (elimination of allosteric inhibition, constitutive product formation); (3) Auxotrophs: products formed via branching biosynthetic pathways such as amino acids and nucleotides
 Antibiotic-resistant mutants: genetic marker / increased cell permeability / protein synthesis with a higher turnover; (2) Antimetabolite-resistant mutants: analog-resistant mutants can form an excess of metabolites, in some cases through changed regulatory mechanisms (elimination of allosteric inhibition, constitutive product formation); (3) Auxotrophs: products formed via branching biosynthetic pathways such as amino acids and nucleotides.")
28
Strategies for the genetic engineering
Source of DNA Vectors Hosts Metabolic engineering

29
Source of DNA for the genetic engineering of bacteria
Donor chromosomal digest with restriction enzymes Complimentary DNA (cDNA) with addition of linkers Synthetic DNA
 with addition of linkers. Synthetic DNA.")
30
Vectors for the genetic engineering of bacteria
Plasmids: copy number, stability, selection markers, expression vectors (strong promoter and ribosome binding, regulatory switch) Bacteriophages: useful cloning vectors, cosmid
 Bacteriophages: useful cloning vectors, cosmid.")
31
Plasmid stability for the genetic engineering of bacteria
Structural instability: deletion and rearrangements of recombinant plasmids / careful construction of the plasmid and the placement of essential genes within it Segregational instability: complete loss of a plasmid / careful construction of the plasmid and the placement of essential genes within it; maintained under continual selection; constructing so-called suicidal strains that require specific markers on the plasmid for survival (strains are constructed with a lethal marker in the chromosome and a repressor of this marker is located on the plasmid) Integration of a gene(s) into the chromosome is normally the best solution
 Integration of a gene(s) into the chromosome is normally the best solution.")
32
Prokaryotic hosts for the genetic engineering
Generally, G(-) bacteria are able to express genes from G(+) bacteria. However, the converse is not always as readily achievable. Secretion problems: inclusion body, periplasmic space in G(-) bacteria, leader peptide (signal sequence). Cell-wall-less bacteria (L-forms) or G(+) bacteria Instability of certain heterologous proteins within the host (proteases) Incorrect folding of the polypeptide that generates an inactive molecule, which may accumulate to form inclusion bodies within the cell Difficulties in achieving post-translational modification of proteins, such as cleavage, glycosylation or amidation
 bacteria are able to express genes from G(+) bacteria. However, the converse is not always as readily achievable. Secretion problems: inclusion body, periplasmic space in G(-) bacteria, leader peptide (signal sequence). Cell-wall-less bacteria (L-forms) or G(+) bacteria. Instability of certain heterologous proteins within the host (proteases) Incorrect folding of the polypeptide that generates an inactive molecule, which may accumulate to form inclusion bodies within the cell. Difficulties in achieving post-translational modification of proteins, such as cleavage, glycosylation or amidation.")
33
Eukaryotic hosts for the genetic engineering
Saccharomyces cerevisiae (safe, vast amount of information, easily and cheaply grown on an industrial scale, relatively rapid growth rate and readily undergoes genetic manipulation / product yields are relatively low at 1-5% of total protein and some proteins are retained within the periplasm); Methylotrophic Pichia angusta (formerly Hansenula polymorpha) and Pichia pastoris (have strong inducible promoters, post-translational modifications similar to those performed by human cells, do not secrete many of their own proteins into the medium)
; Methylotrophic Pichia angusta (formerly Hansenula polymorpha) and Pichia pastoris (have strong inducible promoters, post-translational modifications similar to those performed by human cells, do not secrete many of their own proteins into the medium)")
34
Metabolic engineering
Complete sequencing of many microbial genomes of food grade organisms Relatively minor metabolic engineering has already been implemented to improve the production of existing metabolites, allow the production of new metabolites, impart new catabolic activities and improve fermentation performance Whole metabolic networks within a microorganism may be restructured and such extensive metabolic engineering has major implications within industrial microbiology

35
平時作業 5 (10%) 尋找一篇有關遺傳工程改良的工業微生物的文章。 重點在於改良目標的獨特性、遺傳操作或篩選方法的設計及改良成果的表現。
紀錄搜尋方法及過程,例如:使用的資料庫名稱、關鍵詞、蒐尋到文章數目等。 於第十六週課堂上繳交論文全文。將重點製成投影片,進行口頭報告。每人報告時間5-8分鐘。

36
問題 篩選到某種有用代謝產物的生產菌株之後,常需要進一步改良菌株以便在經濟可行條件下大量生產。請問有哪些主要的改良標的?
使用突變劑處理微生物之後,常常需要培養一段時間再分離純化突變株。請問其原因為何? 使用E. coli或Pichia pastoris作為生產基因重組蛋白質的宿主,各有何優缺點? 下列名詞的內涵:Antimetabolite-resistant mutants、Directed selection of mutants、Metabolic engineering、Protoplast fusion、 Suicidal strains

Similar presentations






 (UMST) M.Sc. Stud. (Immunology ) Institute of Tropical Medicine.(SAS)>")










 Lecture 10 Microbial Genetics (Text Chapter: 10.1-10.4; 10.6-10.14)>")


 Lecture 11 Biotechnology (Text Chapters: 10.15-10.17; 31.1-31.10)>")


