Download presentation
Presentation is loading. Please wait.

1
Sex determination in cucumber Anandkumar Surendrarao VC221: Vegetable crop breeding May 10, 2006

2
Perfect flowers or Hermaphroditic flowers Both male and female reproductive parts are present on the same flower

3
Perfect flowers or Hermaphroditic flowers Both male and female reproductive parts are present on the same flower

4
Monoecious plants – Imperfect flowers Separate male and female flowers are present on the same plant

5
Dioecious plants – Imperfect flowers Male and female flowers are present on individual separate plants

6
Dioecious plants – Imperfect flowers Male and female flowers are present on individual separate plants

7
Sex determination in cucumber flower

8
ABC model of floral development

9
Paper #1

10
Developmental arrest of whorl 4 in male and whorl 3 in female flowers

11
Cucumber floral MADS box gene expression and sequence Probing female cDNA library with petunia MADS box gene Class C Class B

12
amino-acid conservation amongst MADS box genes CUM1 = Ath AGAMOUS (69%) CUM1 = Antirhinum PLENA (71%) CUM26 = Ath PISTLLATA (69%) CUM26 = Antirhinum GLOBOSA (70%) CUM26 = Petunia FLORAL BINDING PROTEIN 1 (71%)
 CUM1 = Antirhinum PLENA (71%) CUM26 = Ath PISTLLATA (69%) CUM26 = Antirhinum GLOBOSA (70%) CUM26 = Petunia FLORAL BINDING PROTEIN 1 (71%)")
13
In situ hybridization analyses of CUM1 and CUM26 expression in wild type male and female flowers with antisense probe to divergent 3 UTR sequence CUM1 – class C, whorls 3 and 4; CUM26 – class B, whorls 2 and 3 Wild type Expression of homeotic genes is observed even in arrested primordia

14
gp mutant flower phenotypes at 22°C A-D sepal-sepal-flower-X sepal-sepal-X-carpel gp mutant flower phenotypes at 30°C E-J sepal-sepal-carpel-X sepal-sepal-X-carpel

15
In situ hybridization analyses of CUM1 and CUM26 expression in gp mutant male flowers gp mutant CUM26 = GP = class B mutant CUM1 – class C, whorls 3 and 4; CUM26 – class B, whorls 2 and 3 22°C 35°C 22°C 35°C

16
CUM1 hypermorph (over-expression) CUM1 hypomorph (co-suppression) Unisexual to bisexual floral conversion
 CUM1 hypomorph (co-suppression) Unisexual to bisexual floral conversion")
17
ABC model of floral development

18
Selective repression of male or female reproductive organs depends on floral whorl position rather than organ identity

19
Paper #2

20
Genetic and environmental control of cucumber sex determination Genotypes Gynoecious-F-M-- Andromonoecious-ffmm- and Monoecious-ffM-- and Hermaphrodite-F-mm- Ethylene and ethephon – induction of flowers AVG and AgNO 3 – induction of flowers

21
Sex of different cultivars used in this study

22
Development of flower buds in gynoecious cucumber plants

24
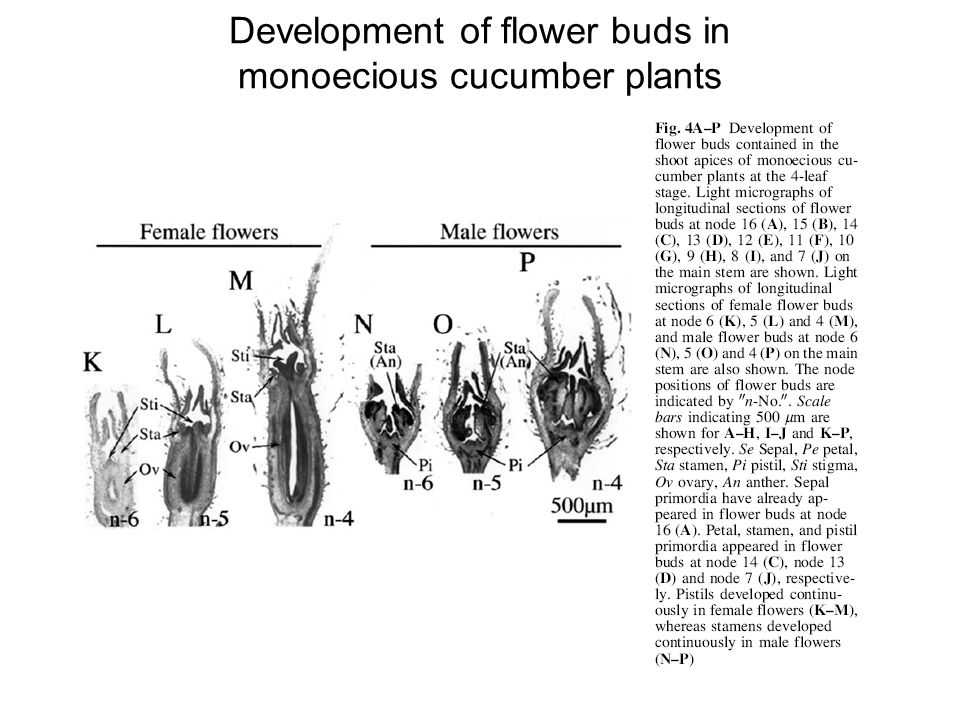
Development of flower buds in monoecious cucumber plants

26
AVG masculinizes between node 8 and 13, Ethephon feminizes between nodes 10 and 14 Floral stages immediately before and after differentiation of stamen primordia are responsive to both AVG and ethephon treatments

27
MonoeciousGynoecious Andromonoecious Monoecious Antisense CS ACS2 Antisense CS ERS Antisense CS ETR1 Antisense CS ETR2 Sense CS ACS2 Sense CS ERS Sense CS ETR1 Sense CS ETR2

28
In situ hybridization results

29
The expression patterns for CS-ACS2, CS-ERS, CS-ETR1, and CS-ETR2 are all different among monoecious, gynoecious and andromonoecious plants. CS-ACS2 and CS-ETR2 are expressed in identical domains in monoecious plants and overlapping domains in gynoecious plants. In andromonoecious plants, none of the ethylene receptors transcripts accumulated in the stamen primordia. Atleast one ethylene receptor transcript is expressed in the stamen and pistil primordia of monoecious and gynoecious flowers, and pistil primordium of andromonoecious flowers.

30
Cells producing and sensing ethylene are identical. Eg. Overlapping CS-ACS2 and CS-ETR2 mRNA expression in monoecious and gynoecious plants, direct determination of female flowers by inducing pistil development. Cells producing and sensing ethylene are adjacent. Eg. mRNA expression of CS-ACS2 in adaxial side of petals but all the receptors in stamen primordia in monoecious plants. (diffusion?) Cells producing and sensing ethylene are distant. Eg. mRNA expression of CS-ACS2 in pistil primordia but that of receptors in the stamen primordia. (diffusion?)
 Cells producing and sensing ethylene are distant. Eg. mRNA expression of CS-ACS2 in pistil primordia but that of receptors in the stamen primordia. (diffusion ).")
31
Paper #3

32
What are the downstream targets of the sex determination machinery that allow the selective arrest of stamen and pistil primordia development? Use suppression subtractive hybridization on NILs of gynoecious (FFMMaa), hermaphrodite (FFmmaa), androecious (ffMMaa) and monoecious (ffMMA-) genotypes. AgNO 3 induced male flowers in gynoecious plants, and ethephon induced female flowers in hermaphrodite plants used for SSH. Controls for SSH were female and male flowers from gynoecious and androecious plants respectively.
, hermaphrodite (FFmmaa), androecious (ffMMaa) and monoecious (ffMMA-) genotypes. AgNO 3 induced male flowers in gynoecious plants, and ethephon induced female flowers in hermaphrodite plants used for SSH. Controls for SSH were female and male flowers from gynoecious and androecious plants respectively..")
33
Results from SSH

34
Selection of 21/178 clones by dot blot analyses 11/21 differentially expressed in hermaphrodite buds – Clone #38 is putative CS nt sugar epimerase 10/21 differentially expressed in gynoecious plants

35
Putative sugar nt epimerase expressed lower in gynoecious than in hermaphrodite plants Floral buds Leaves

36
Putative sugat nt epimerase expressed higher in natural/induced male flowers compared to natural/induced female flowers Monoecious + no treatment plants + ethephon plants + AgNO 3

37
No detectable polymorphisms at gDNA level between gynoecious and hermaphrodite plants Southern blot hybridization with 19 different restriction enzymes

38
Mechanistic role for sugar nt epimerase in stamen primordia outgrowth and arrest UDP glucose-4-epimerase converts UDP-glucose to UDP- galactose. These are required for the synthesis of AGPs (Arabino- Galactan proteins) and cell wall polysaccharides that are necessary for cell wall expansion and therefore primordial outgrowth.
 and cell wall polysaccharides that are necessary for cell wall expansion and therefore primordial outgrowth..")
Similar presentations











 have two growth phases – vegetative growth, production of stems and leaves, occurs at the apical meristem,>")






 Examine the expression of the gene (GUS, In situ)>")
 A, B, C, D FRUITFULL,>")
