Download presentation
Presentation is loading. Please wait.

1
Репликация, транскрипция, генетичен код, транслация
Атанаска Андреева, катедра “Физика на конд. материя” Физически факултет, СУ “Св. Кл. Охридски” бул. “Дж. Баучър” 5, 1164 София

2
Хромозоми

3
Репликация на ДНК - анимация

4
Транскрипция - презапис
Translation: DNA to mRNA to Protein the decoding of instructions for making proteins, involving mRNA in transcription as well as tRNA. two major steps. In the first step, the information in DNA is transferred to a messenger RNA (mRNA) molecule by way of a process called transcription. During transcription, the DNA of a gene serves as a template for complementary base-pairing, and an enzyme called RNA polymerase III catalyzes the formation of a pre-mRNA molecule, which is then processed to form mature mRNA The resulting mRNA is a single-stranded copy of the gene, which next must be translated into a protein molecule.
 molecule by way of a process called transcription. During transcription, the DNA of a gene serves as a template for complementary base-pairing, and an enzyme called RNA polymerase III catalyzes the formation of a pre-mRNA molecule, which is then processed to form mature mRNA The resulting mRNA is a single-stranded copy of the gene, which next must be translated into a protein molecule.")
5
Транскрипция Анимация на транскрипция запис:

6
Генетичен код During translation, which is the second major step in gene expression, the mRNA is "read" according to the genetic code, which relates the DNA sequence to the amino acid sequence in proteins. Кодовото съотношение е: 4 букви (НК) и 20 букви (а.к.): 42=16<20, но 43=64>20 (Гамов) Кодон – три бази, кодиращи 1 а.к. Намерени са експериментално (Ниренберг, Корана)
 и 20 букви (а.к.): 42=16<20, но 43=64>20 (Гамов) Кодон – три бази, кодиращи 1 а.к. Намерени са експериментално (Ниренберг, Корана)")
7
Кодони 64 codons One codon: Met, Trp.
Two codons: Asn, Asp, Cys, Gln, Glu, His, Lys, Phe, Tyr, Three codons: Ile, STOP ("nonsense"). Four codons: Ala, Gly, Pro, Thr, Val. Five codons: none. Six codons: Arg, Leu, Ser
. Four codons: Ala, Gly, Pro, Thr, Val. Five codons: none. Six codons: Arg, Leu, Ser.")
8
Рибозома Where Translation Occurs
Within all cells, the translation machinery resides within a specialized organelle called the ribosome. In eukaryotes, mature mRNA molecules must leave the nucleus and travel to the cytoplasm, where the ribosomes are located. On the other hand, in prokaryotic organisms, ribosomes can attach to mRNA while it is still being transcribed. In this situation, translation begins at the 5' end of the mRNA while the 3' end is still attached to DNA. In all types of cells, the ribosome is composed of two subunits: the large (50S) subunit and the small (30S) subunit (S, for svedberg unit, is a measure of sedimentation velocity and, therefore, mass). Each subunit exists separately in the cytoplasm, but the two join together on the mRNA molecule. The ribosomal subunits contain proteins and specialized RNA molecules—specifically, ribosomal RNA (rRNA) and transfer RNA (tRNA). The tRNA molecules are adaptor molecules—they have one end that can read the triplet code in the mRNA through complementary base-pairing, and another end that attaches to a specific amino acid (Chapeville et al., 1962; Grunberger et al., 1969). The idea that tRNA was an adaptor molecule was first proposed by Francis Crick, co-discoverer of DNA structure, who did much of the key work in deciphering the genetic code (Crick, 1958). Within the ribosome, the mRNA and aminoacyl-tRNA complexes are held together closely, which facilitates base-pairing. The rRNA catalyzes the attachment of each new amino acid to the growing chain.
 subunit and the small (30S) subunit (S, for svedberg unit, is a measure of sedimentation velocity and, therefore, mass). Each subunit exists separately in the cytoplasm, but the two join together on the mRNA molecule. The ribosomal subunits contain proteins and specialized RNA molecules—specifically, ribosomal RNA (rRNA) and transfer RNA (tRNA). The tRNA molecules are adaptor molecules—they have one end that can read the triplet code in the mRNA through complementary base-pairing, and another end that attaches to a specific amino acid (Chapeville et al., 1962; Grunberger et al., 1969). The idea that tRNA was an adaptor molecule was first proposed by Francis Crick, co-discoverer of DNA structure, who did much of the key work in deciphering the genetic code (Crick, 1958). Within the ribosome, the mRNA and aminoacyl-tRNA complexes are held together closely, which facilitates base-pairing. The rRNA catalyzes the attachment of each new amino acid to the growing chain.")
9
Превод – инициация: синтез на първата а.к.
Translation Begins After the Assembly of a Complex Structure

10
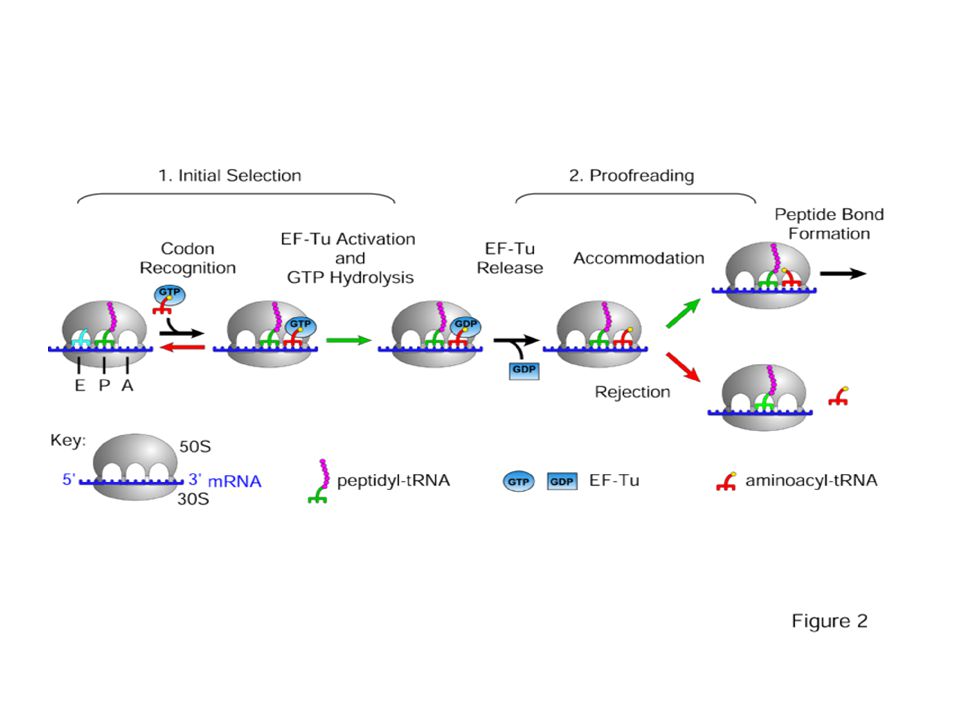
Превод - елонгация The Elongation Phase

11
The Elongation Phase The next phase in translation is known as the elongation phase (Figure 5). First, the ribosome moves along the mRNA in the 5'-to-3'direction, which requires the elongation factor G, in a process called translocation. The tRNA that corresponds to the second codon can then bind to the A site, a step that requires elongation factors (in E. coli, these are called EF-Tu and EF-Ts), as well as guanosine triphosphate (GTP) as an energy source for the process. Upon binding of the tRNA-amino acid complex in the A site, GTP is cleaved to form guanosine diphosphate (GDP), then released along with EF-Tu to be recycled by EF-Ts for the next round.
. First, the ribosome moves along the mRNA in the 5 -to-3 direction, which requires the elongation factor G, in a process called translocation. The tRNA that corresponds to the second codon can then bind to the A site, a step that requires elongation factors (in E. coli, these are called EF-Tu and EF-Ts), as well as guanosine triphosphate (GTP) as an energy source for the process. Upon binding of the tRNA-amino acid complex in the A site, GTP is cleaved to form guanosine diphosphate (GDP), then released along with EF-Tu to be recycled by EF-Ts for the next round.")
12
The Elongation Phase Next, peptide bonds between the now-adjacent first and second amino acids are formed through a peptidyl transferase activity. For many years, it was thought that an enzyme catalyzed this step, but recent evidence indicates that the transferase activity is a catalytic function of rRNA (Pierce, 2000). After the peptide bond is formed, the ribosome shifts, or translocates, again, thus causing the tRNA to occupy the E site. The tRNA is then released to the cytoplasm to pick up another amino acid. In addition, the A site is now empty and ready to receive the tRNA for the next codon. This process is repeated until all the codons in the mRNA have been read by tRNA molecules, and the amino acids attached to the tRNAs have been linked together in the growing polypeptide chain in the appropriate order. At this point, translation must be terminated, and the nascent protein must be released from the mRNA and ribosome. Termination of Translation There are three termination codons that are employed at the end of a protein-coding sequence in mRNA: UAA, UAG, and UGA. No tRNAs recognize these codons. Thus, in the place of these tRNAs, one of several proteins, called release factors, binds and facilitates release of the mRNA from the ribosome and subsequent dissociation of the ribosome.
. After the peptide bond is formed, the ribosome shifts, or translocates, again, thus causing the tRNA to occupy the E site. The tRNA is then released to the cytoplasm to pick up another amino acid. In addition, the A site is now empty and ready to receive the tRNA for the next codon. This process is repeated until all the codons in the mRNA have been read by tRNA molecules, and the amino acids attached to the tRNAs have been linked together in the growing polypeptide chain in the appropriate order. At this point, translation must be terminated, and the nascent protein must be released from the mRNA and ribosome. Termination of Translation. There are three termination codons that are employed at the end of a protein-coding sequence in mRNA: UAA, UAG, and UGA. No tRNAs recognize these codons. Thus, in the place of these tRNAs, one of several proteins, called release factors, binds and facilitates release of the mRNA from the ribosome and subsequent dissociation of the ribosome.")
14
[E,P,A] three tRNA binding sites on 30S subunit
![[E,P,A] three tRNA binding sites on 30S subunit](http://slideplayer.com/slide/1665275/7/images/14/%5BE%2CP%2CA%5D+three+tRNA+binding+sites+on+30S+subunit.jpg "[E,P,A] three tRNA binding sites on 30S subunit")
15
ДНК-РНК-Б – централната догма на МБ
РНК’-ДНК-РНК-Б – обратна транскрипция Анимация на процеса превод

16
Нобеловите награди по химия за 2009 - рибозомата
Nobel for explaining nature's protein factories This year's Nobel Prize in Chemistry awards Venkatraman Ramakrishnan, Thomas A. Steitz and Ada E. Yonath for having showed what the ribosome looks like and how it functions at the atomic level. All three have used a method called X-ray crystallography to map the position for each and every one of the hundreds of thousands of atoms that make up the ribosome.

17
Venkatraman Ramakrishnan, US citizen
Venkatraman Ramakrishnan, US citizen. Born in 1952 in Chidambaram, Tamil Nadu, India. Ph.D. in Physics in 1976 from Ohio University, USA. Senior Scientist and Group Leader at Structural Studies Division, MRC Laboratory of Molecular Biology, Cambridge, UK. 30S subunit, view of the intersubunit interface 30S subunit, back view (1462x1745)
")
18
Induced fit and codon-anticodon recognition on tRNA binding (avi format, 52 Mb)
A few years ago, we determined the complete atomic structure of the 30S subunit and its complexes with several antibiotics, initiation factor IF1, and cognate and near-cognate tRNA anticodon stem-loops complexed with mRNA in the A site. More recently, we have determined the high-resolution structure of the entire ribosome complexed with mRNA and tRNA. These studies have shed light on antibiotic function, and the mechanism of tRNA and mRNA recognition and decoding by the ribosome. 30S subunit with antibiotics shown as light sources (800x1000)
")
25
Thomas A. Steitz, US citizen. Born in 1940 in Milwaukee, WI, USA. Ph.D. in Molecular Biology and Biochemistry in 1966 from Harvard University, MA, USA. Sterling Professor of Molecular Biophysics and Biochemistry and Howard Hughes Medical Institute Investigator, both at Yale University, CT, USA. Our recent accomplishments have included the determination of the atomic structure of the 50S ribosomal subunit and its complexes with substrate, intermediate and product analogues as well as complexes with more than a dozen antibiotics. These structures establish that the ribosome is a ribozyme, provide insights into the mechanism of peptide bond formation and show how several classes of antibiotics function. In the area of transcription, six structures of T7 RNA polymerase captured in various functional states show the structural basis of the transition from the initiation to elongation phase, which involves a large protein structural rearrangement. They explain the basis of promoter clearance, processivity of the elongation phase, translocation and strand separation. The structures of the CCA-adding enzyme captured in each state of CCA incorporation onto tRNA explain the enzyme's specificity for nucleotide incorporation in the absence of a nucleic acid template. The structure of a recombination intermediate of γδ resolvase suggests that site specific recombination by this enzyme is achieved by subunit rotation.

26
Prof Ada Yonath: Structural Biology Department, Weizmann Institute of Science, Rehovot, Israel

Similar presentations























 of polypeptide chains (proteins) Two phases: Transcription & Translation mRNA must be processed before it.>")

