Download presentation
Presentation is loading. Please wait.

1
Major Histocompatibility Complex

2
Antigen Presentation APC to T-cell

3
MHC The immune system relies on many regulatory mechanisms that govern its ability to respond to infectious agents and neoplastic tissues, but no single scheme is as much a cellular and molecular microcosm of complex biologic systems as that controlled by the Major Histocompatibility Complex (Mhc).
.")
4
MHC The Mhc is a set of linked genes, located on chromosome 6 of the human, chromosome 17 of the mouse, and chromosome 20 of the rat, that was first identified for its effects on tumor or skin transplantation and control of immune responsiveness.

5
MHC Although the control of transplantation, autoimmunity, and the other immune responses are the phenotypic consequences of the function of molecules encoded in the Mhc, understanding the Mhc becomes clear if we think of it in molecular and cellular terms. MHC molecules are cell surface receptors that bind antigen fragments and display them to various cells of the immune system, most importantly T-cells that bear ab receptors.

6
MHC The goal of this lecture will be to outline the general principles of molecular organization and function of both the genetic regions that encode MHC molecules and the functional cell surface molecules themselves.

7
Major Histocompatibility Complex
The major function of the molecules encoded by the MHC is to facilitate the display of unique molecular fragments on the surface of cells in an arrangement that permits their recognition by immune effectors such as T-lymphocytes.

8
Major Histocompatibility Complex
The MHC molecule accomplishes its major role in immune recognition by satisfying two distinct molecular functions: the binding of peptides (or in some cases nonpeptidic molecules) and the interaction with T cells, usually via the ab T-cell receptor (TCR).
 and the interaction with T cells, usually via the ab T-cell receptor (TCR).")
9
Major Histocompatibility Complex
The binding of peptides by an MHC-I or MHC-II molecule is the selective event that permits the cell expressing the MHC molecule (the antigen-presenting cell, APC) to sample either its own proteins (in the case of MHC-I) or the proteins ingested from the immediate extracellular environment (in the case of MHC-II).
 to sample either its own proteins (in the case of MHC-I) or the proteins ingested from the immediate extracellular environment (in the case of MHC-II).")
10
MHC I In particular, cell surface MHC class I glycoproteins gather from the cell’s biosynthetic pathway fragments of proteins derived from infecting viruses, intracellular parasites, or self molecules, either normally expressed or dysregulated by tumorigenesis, and then display these molecular fragments at the cell surface.

11
Fig 4-4

12
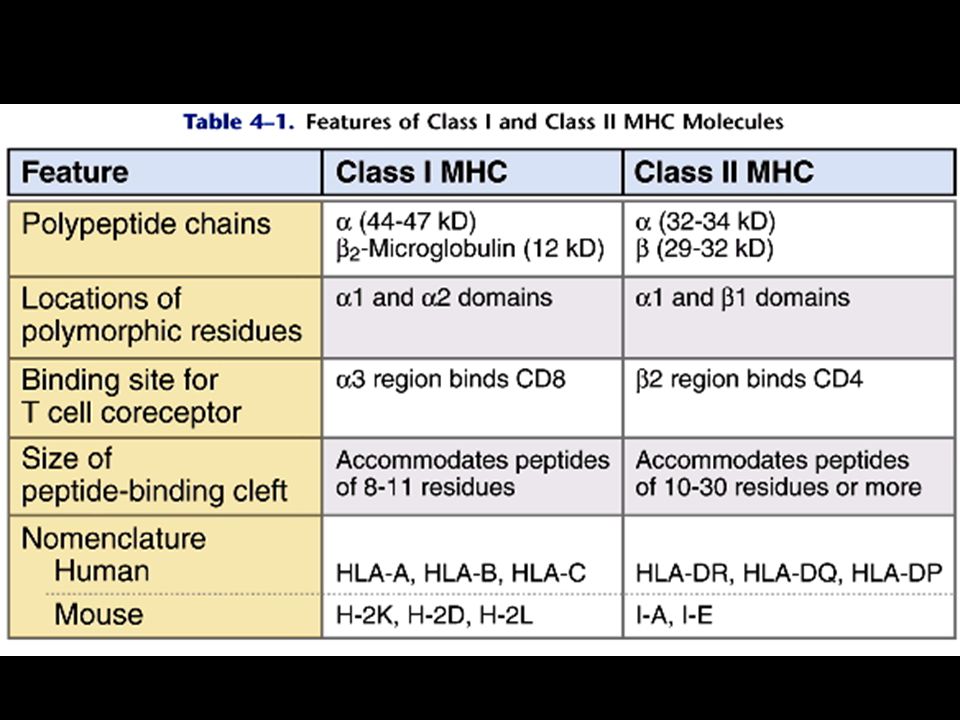
MHC Class I The a chains forms a platform of 8-stranded anti-parallel b pleated sheet supporting two parallel strands of a-helix. The formed cleft can bind peptides of amino acids in a flexible extended conformation.

13
MHC Class I The a3 segment of the MHC I serves as a binding site for CD8. The b2-microglobulin interacts with the a3 non-covalently.

14
MHC Class I The a chain the HLA-A, -B, -C

17
MHC Class II Class II are composed of 2 non-covalent associated peptides, a chain and b chain. The peptide binding cleft can accommodate peptides up to 30 amino acids in length.

18
MHC Class II The b2 is the binding site for the CD4.
The a chains are the HLA DR The b chains are the HLA DQ and DP

19
MHC Class II

21
MHC I Peptides derived from proteins found in the cytosol that are then degraded by the multiproteolytic proteasome complex into peptides.

22
MHC I The resulting peptides, transported from the cytosol to the endoplasmic reticulum with the aid of the intrinsic membrane transporter, the transporter associated with antigen processing (TAP), are then cooperatively folded into the newly synthesized MHC-I molecule.
, are then cooperatively folded into the newly synthesized MHC-I molecule.")
23
MHC I The MHC-I molecule, the ability to interact with TCR, then allows the APC bearing a particular MHC–peptide complex to engage an appropriate ab TCR as the first step in the activation of a CD8+ cellular program that might lead to cytolysis of the APC as a target and/or to the secretion of lymphokines by the T cell.

24
MHC I Differential response
Potentially any nucleated cell

25
MHC II MHC class II molecules bind peptides derived from the degradation of proteins ingested by MHC-II–expressing APC, and display them at the cell surface for recognition by specific T-lymphocytes.

26
MHC II The MHC-II antigen presentation pathway is based on the initial assembly of the MHC-II ab heterodimer with a dual function molecule.

27
MHC II The invariant chain (Ii) that serves as a chaperone to direct the ab heterodimer to an endosomal, acidic protein–processing location.
 that serves as a chaperone to direct the ab heterodimer to an endosomal, acidic protein–processing location.")
28
MHC II Where Ii encounters antigenic peptides, it serves to protect the antigen-binding site of the MHC-II molecule so that it preferentially will be loaded with antigenic peptides in this endosomal–lysosomal location.

29
MHC II The loading of the MHC-II molecule with antigenic peptide, a process dependent on the release of the Ii-derived CLIP peptide, in part dependent on the MHC-II-like molecule leads to the cell surface expression of MHC-II peptide complexes.

30
MHC II TH 1 TH 2

31
MHC I versus MHC II The MHC-I antigen presentation pathway is most easily thought of as an inside-out pathway by which protein fragments of molecules synthesized by the cell are delivered to and bound by the MHC-I molecule during its biosynthesis. In contrast, the MHC-II antigen presentation pathway is best more clearly visualized as an outside-in one in which ingested proteins are degraded by enzymes in the endosomal–lysosomal system and are delivered to the MHC-II molecules in that degradative compartment.

32
MHC I versus MHC II The MHC-I and MHC-II molecules also show preferential restriction to T cells of the CD8- or CD4-bearing subsets. This is related to the observation that CD8 binds to the nonpolymorphic a3 domain of MHC-I molecules, while CD4 interacts with membrane proximal domains of MHC-II.

33
Viruses and MHC There is little question that the immune system plays a decisive role in host defense against microbial pathogens, and it should not have come as a surprise to find that many pathogens have the ability to produce molecules that modulate the host response.

34
Viruses and MHC The most striking results have been obtained for viruses and MHC class I molecules. (a) peptide binding to TAP (ICP47 of herpes simplex) or peptide translocation by TAP (US6 of human cytomegalovirus); (b) stable insertion of the class I heavy chain into the ER membrane (US2 and US11 of human cytomegalovirus and HIV vpu;
 peptide binding to TAP (ICP47 of herpes simplex) or peptide translocation by TAP (US6 of human cytomegalovirus); (b) stable insertion of the class I heavy chain into the ER membrane (US2 and US11 of human cytomegalovirus and HIV vpu;")
35
Viruses and MHC The most striking results have been obtained for viruses and MHC class I molecules. (c) release of assembled class I–peptide complexes from the ER (E19 of adenovirus); (d) inhibition of MHC class I heavy-chain gene transcription (adenovirus, HIV); and (e) accelerated clearance of surface class I (HIV Nef).
 release of assembled class I–peptide complexes from the ER (E19 of adenovirus); (d) inhibition of MHC class I heavy-chain gene transcription (adenovirus, HIV); and. (e) accelerated clearance of surface class I (HIV Nef).")
36
Viruses and MHC The most striking results have been obtained for viruses and MHC class II molecules. Epstein-Barr virus produces a molecule that binds the human DR b chain. The T-cell response to herpes simplex in humans has been found to have the unusual feature of eliciting primarily CD4+ cytotoxic cells, and not stimulating much in the way of a CD8+ CTL response.

37
Viruses and MHC The most striking results have been obtained for viruses and MHC class II molecules. Cells infected with herpes simplex were found to have a defect in the stable assembly of MHC class I heavy chain–-b2m dimers and the export of class I molecules to the cell surface, which later was recognized to resemble the situation in cells lacking effective peptide import into the ER as a result of mutations in TAP-1 or -2.

38
MHC I versus NK Cells The expression by the target cell of MHC class I molecules can, in certain cases, protect the target from killing by the NK effector events, and target cells defective in the expression of normal MHC class I cells are susceptible to such NK-cell lysis.

39
Human MHC Gene - Chromosome 6
The genes encoding the chains of MHC class I molecules and the a and b chains of MHC class II molecules are linked within the complex; the genes for b2-microglobulin and the invariant chain lie on separate chromosomes. Separate regions contain the genes encoding the MHC class I and MHC class II molecules, and within these regions there are several genes for each chain.

40
Human Leukocyte Antigen (HLA)
In humans, there are three MHC Class I a-chain genes, called: HLA -A, -B, and -C. There are also three pairs of MHC Class II a- and b-chain genes, called HLA-DR, -DP, and -DQ.

41
HLA Tissue Type example: A1,28 B38,44 DR 4, 11

42
Autoimmunity The mechanisms of tissue damage in autoimmune diseases are essentially the same as those that operate in protective immunity. Autoimmune responses are a natural consequence of the open repertoires of both B-cell and T-cell receptor that allows them to recognize any pathogen.

43
Autoimmunity HLA genotype Diabetes Mellitus DR3, DR4
Juvenile Rhematoid Arth. B27 Multiple Sclerosis DR2

44
Mechanisms of Autoimmunity
Another mechanism may by through the binding of a superantigen

45
HLA vs. Transplantation
HLA matching between the donor and recipient - a predictor of graft survival in the kidney allograft patient HLA mis-matches may cause chronic pathologies in the allograft It is clear that a negative HLA-A2 recipient receiving an allograft expressing HLA-A2 will have a much higher risk of humoral rejection of the vascular endothelium.

46
We love learning about MHC. Monday we will Have an exam

Similar presentations




>")





 Anindyaningrum Chrisant Rystiasih Nadia Karlina *Protein images comparing the MHC I (1hsa) and MHC II (1dlh) molecules.>")


 Ganys Tri S. (115090107111020) Agatha Mia(115090101111010) Vita Agustina(115090100111011)>")




 & MHC Complexes-Antigen Presentation>")


